

A Novel Alphabaculovirus from the Soybean Looper, Chrysodeixis includens, that Produces Tetrahedral Occlusion Bodies and Encodes Two Copies of he65
- PMID: 31247912
- PMCID: PMC6669638
- DOI: 10.3390/v11070579
A Novel Alphabaculovirus from the Soybean Looper, Chrysodeixis includens, that Produces Tetrahedral Occlusion Bodies and Encodes Two Copies of he65
Abstract
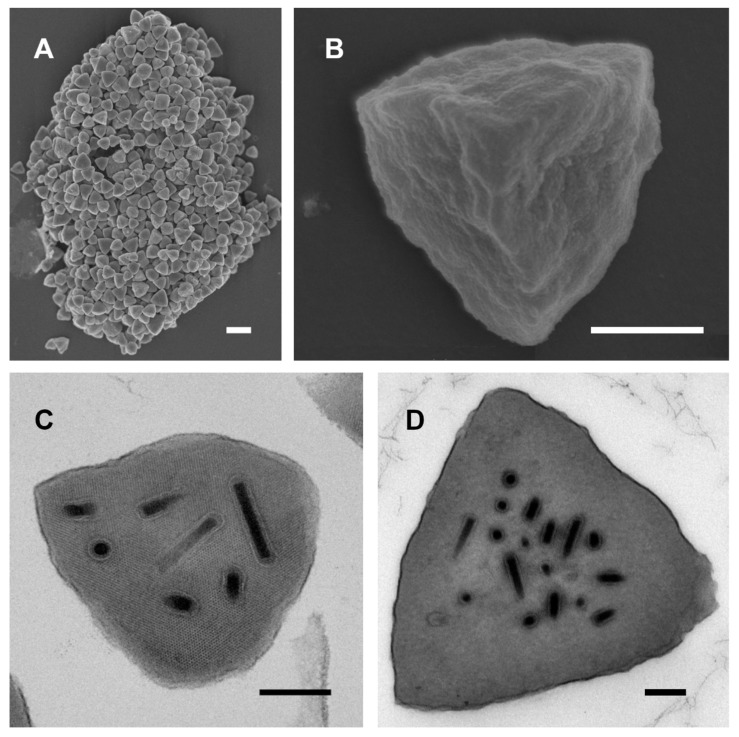
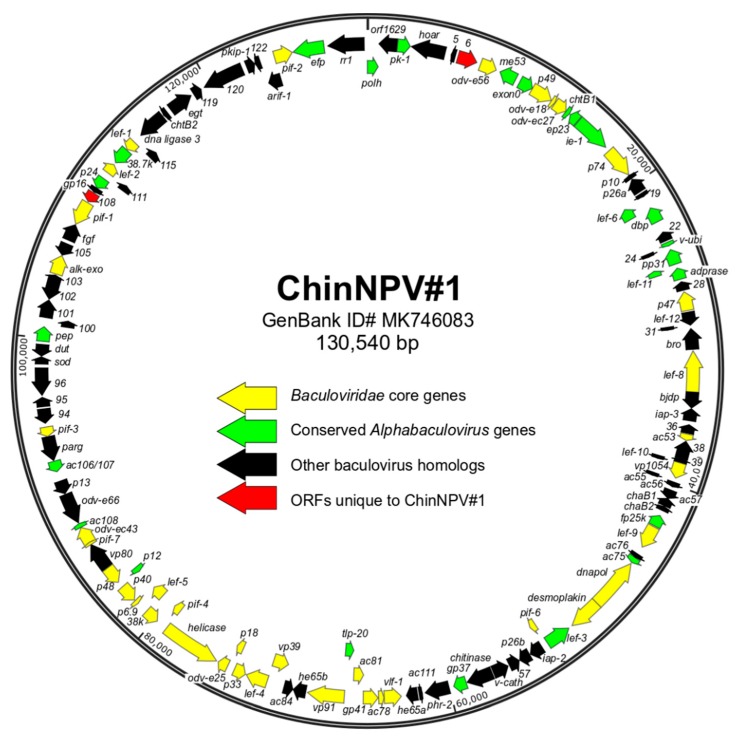
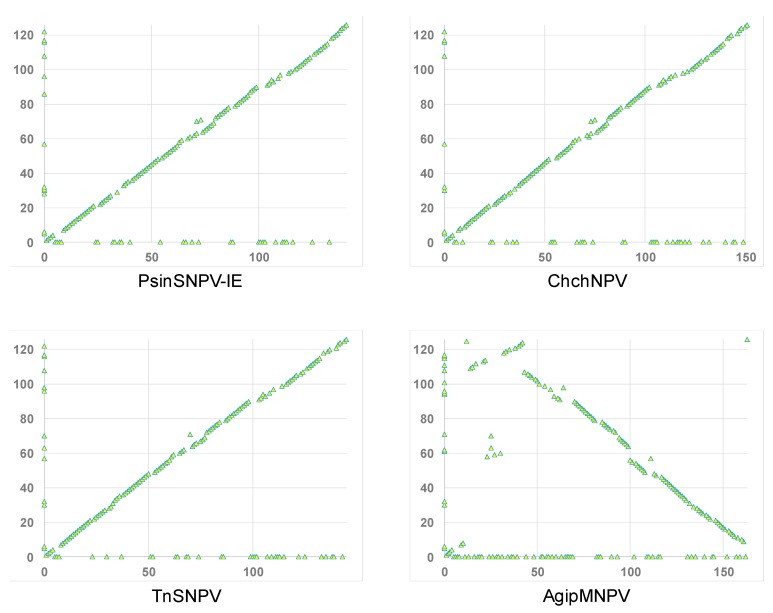
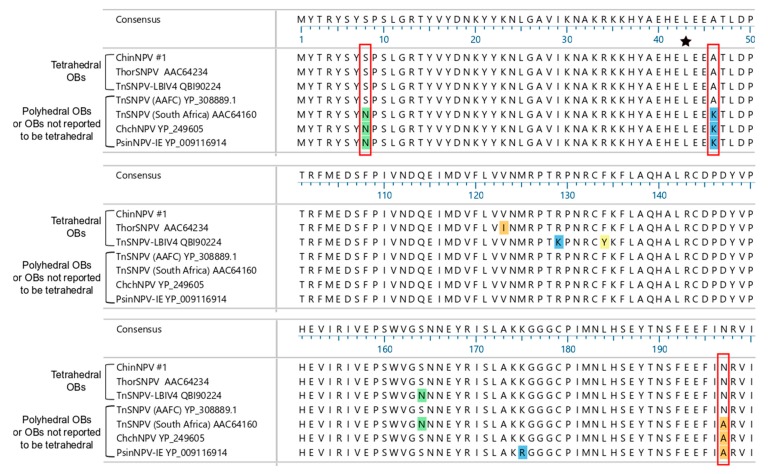
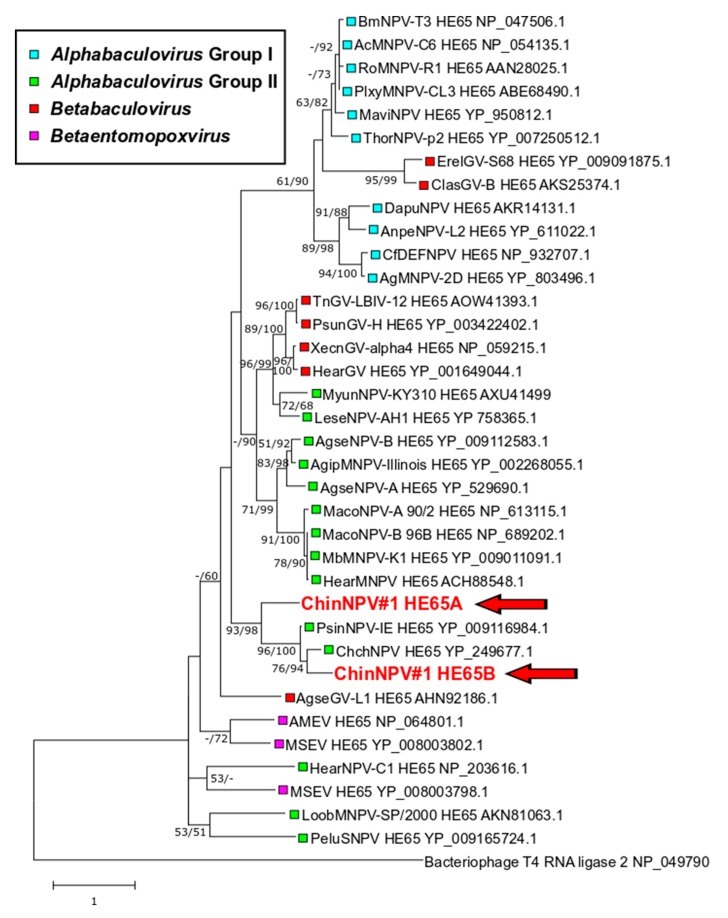
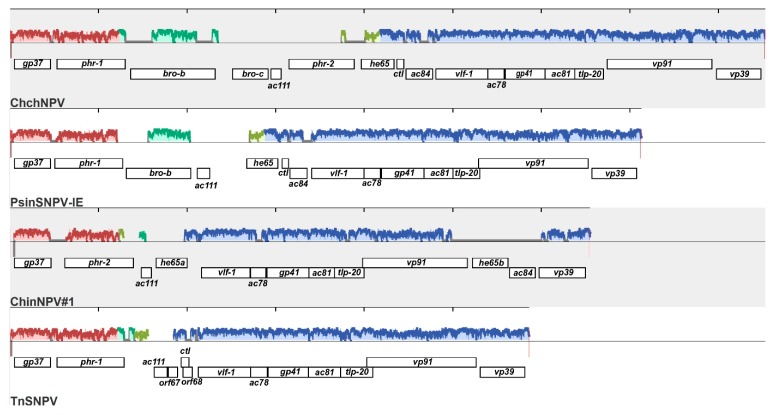
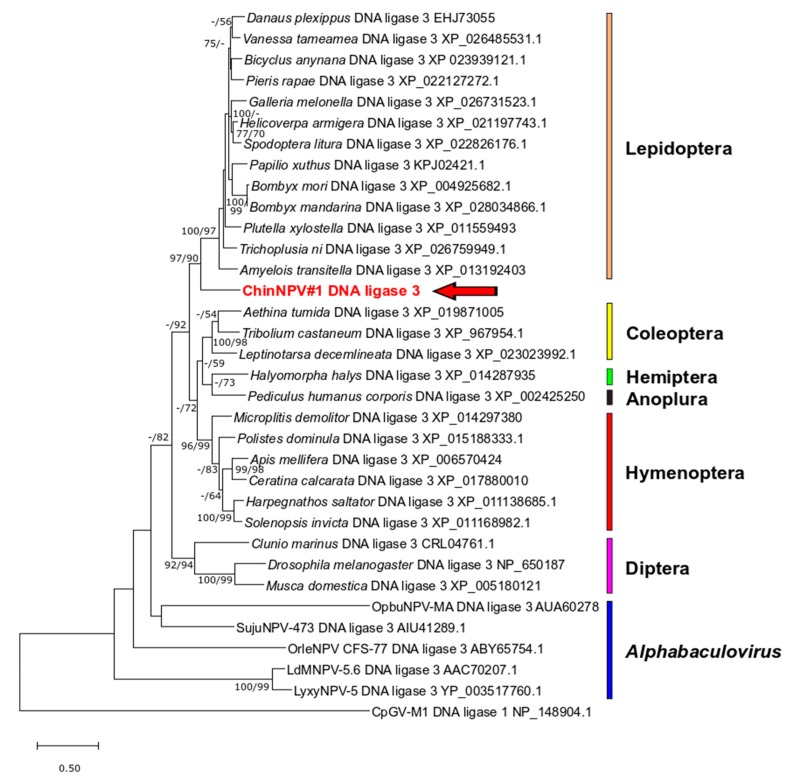
Isolates of the alphabaculovirus species, Chrysodeixis includens nucleopolyhedrovirus, have been identified that produce polyhedral occlusion bodies and infect larvae of the soybean looper, Chrysodeixis includens. In this study, we report the discovery and characterization of a novel C. includens-infecting alphabaculovirus, Chrysodeixis includens nucleopolyhedrovirus #1 (ChinNPV#1), that produces tetrahedral occlusion bodies. In bioassays against C. includens larvae, ChinNPV #1 exhibited a degree of pathogenicity that was similar to that of other ChinNPV isolates, but killed larvae more slowly. The host range of ChinNPV#1 was found to be very narrow, with no indication of infection occurring in larvae of Trichoplusia ni and six other noctuid species. The ChinNPV#1 genome sequence was determined to be 130,540 bp, with 126 open reading frames (ORFs) annotated but containing no homologous repeat (hr) regions. Phylogenetic analysis placed ChinNPV#1 in a clade with other Group II alphabaculoviruses from hosts of lepidopteran subfamily Plusiinae, including Chrysodeixis chalcites nucleopolyhedrovirus and Trichoplusia ni single nucleopolyhedrovirus. A unique feature of the ChinNPV#1 genome was the presence of two full-length copies of the he65 ORF. The results indicate that ChinNPV#1 is related to, but distinct from, other ChinNPV isolates.
Keywords: Chrysodeixis includens; DNA ligase 3; baculovirus; he65; lef-12; occlusion body; polyhedrin; soybean looper.
Conflict of interest statement
The authors declare no conflict of interest. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture. USDA is an equal opportunity provider and employer.
Figures


Similar articles
-
Mosaic genome evolution and phylogenetics of Chrysodeixis includens nucleopolyhedrovirus (ChinNPV) and virulence of seven new isolates from the Brazilian states of Minas Gerais and Mato Grosso.Arch Virol. 2021 Jan;166(1):125-138. doi: 10.1007/s00705-020-04858-2. Epub 2020 Oct 27. Arch Virol. 2021. PMID: 33111162
-
No cross-resistance between ChinNPV and chemical insecticides in Chrysodeixis includens (Lepidoptera: Noctuidae).J Invertebr Pathol. 2019 Jun;164:66-68. doi: 10.1016/j.jip.2019.05.001. Epub 2019 May 9. J Invertebr Pathol. 2019. PMID: 31078547
-
Genetic Variability of Chrysodeixis Includens Nucleopolyhedrovirus (ChinNPV) and the Insecticidal Characteristics of Selected Genotypic Variants.Viruses. 2019 Jun 26;11(7):581. doi: 10.3390/v11070581. Viruses. 2019. PMID: 31247955 Free PMC article.
-
The complete genome of Rachiplusia nu nucleopolyhedrovirus (RanuNPV) and the identification of a baculoviral CPD-photolyase homolog.Virology. 2019 Aug;534:64-71. doi: 10.1016/j.virol.2019.05.019. Epub 2019 May 29. Virology. 2019. PMID: 31200103
-
Generation of Variability in Chrysodeixis includens Nucleopolyhedrovirus (ChinNPV): The Role of a Single Variant.Viruses. 2021 Sep 22;13(10):1895. doi: 10.3390/v13101895. Viruses. 2021. PMID: 34696324 Free PMC article.
Cited by
-
Ecological interactions, host plant defenses, and control strategies in managing soybean looper, Chrysodeixis includens (Lepidoptera: Noctuidae).Front Insect Sci. 2024 Dec 12;4:1480940. doi: 10.3389/finsc.2024.1480940. eCollection 2024. Front Insect Sci. 2024. PMID: 39726917 Free PMC article. Review.
-
Complete Genome Sequences of Seven New Chrysodeixis includens Nucleopolyhedrovirus Isolates from Minas Gerais and Mato Grosso States in Brazil.Microbiol Resour Announc. 2020 Feb 20;9(8):e01501-19. doi: 10.1128/MRA.01501-19. Microbiol Resour Announc. 2020. PMID: 32079633 Free PMC article.
References
-
- Eichlin T.D., Cunningham H.B. The Plusiinae (Lepidoptera: Noctuidae) of America north of Mexico, Emphasizing Genitalic and Larval Morphology. United States Department of Agriculture, Agricultural Research Service; Washington, DC, USA: 1978. p. 122.
-
- Kogan M., Turnipseed S.G. Ecology and management of soybean arthropods. Ann. Rev. Entomol. 1987;32:507–538. doi: 10.1146/annurev.en.32.010187.002451. - DOI
-
- Bortolotto O.C., de F. Bueno R.C.O., de F. Bueno A., da Kruz A.K.S., Queiroz A.P., Sanzovo A., Ferreira R.B. The use of soybean integrated pest management in Brazil: A review. Agron. Sci. Biotechnol. 2015;1:25–32.
-
- Owen L.N., Catchot A.L., Musser F.R., Gore J., Cook D.C., Jackson R. Susceptibility of Chrysodeixis includens (Lepidoptera:Noctuidae) to reduced-risk insecticides. Fla. Entomol. 2013;96:554–559. doi: 10.1653/024.096.0221. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
