

Enzymes for Detoxification of Various Mycotoxins: Origins and Mechanisms of Catalytic Action
- PMID: 31247992
- PMCID: PMC6651818
- DOI: 10.3390/molecules24132362
Enzymes for Detoxification of Various Mycotoxins: Origins and Mechanisms of Catalytic Action
Abstract
Mycotoxins are highly dangerous natural compounds produced by various fungi. Enzymatic transformation seems to be the most promising method for detoxification of mycotoxins. This review summarizes current information on enzymes of different classes to convert various mycotoxins. An in-depth analysis of 11 key enzyme mechanisms towards dozens of major mycotoxins was realized. Additionally, molecular docking of mycotoxins to enzymes' active centers was carried out to clarify some of these catalytic mechanisms. Analyzing protein homologues from various organisms (plants, animals, fungi, and bacteria), the prevalence and availability of natural sources of active biocatalysts with a high practical potential is discussed. The importance of multifunctional enzyme combinations for detoxification of mycotoxins is posed.
Keywords: antidote; biochemical mechanism; conversion; detoxification; enzyme; molecular modeling; mycotoxin; origins.
Conflict of interest statement
EE and IL are named co-inventors of several Russian patents that involve organophosphorus hydrolase and its modified forms as a base active compound.
Figures
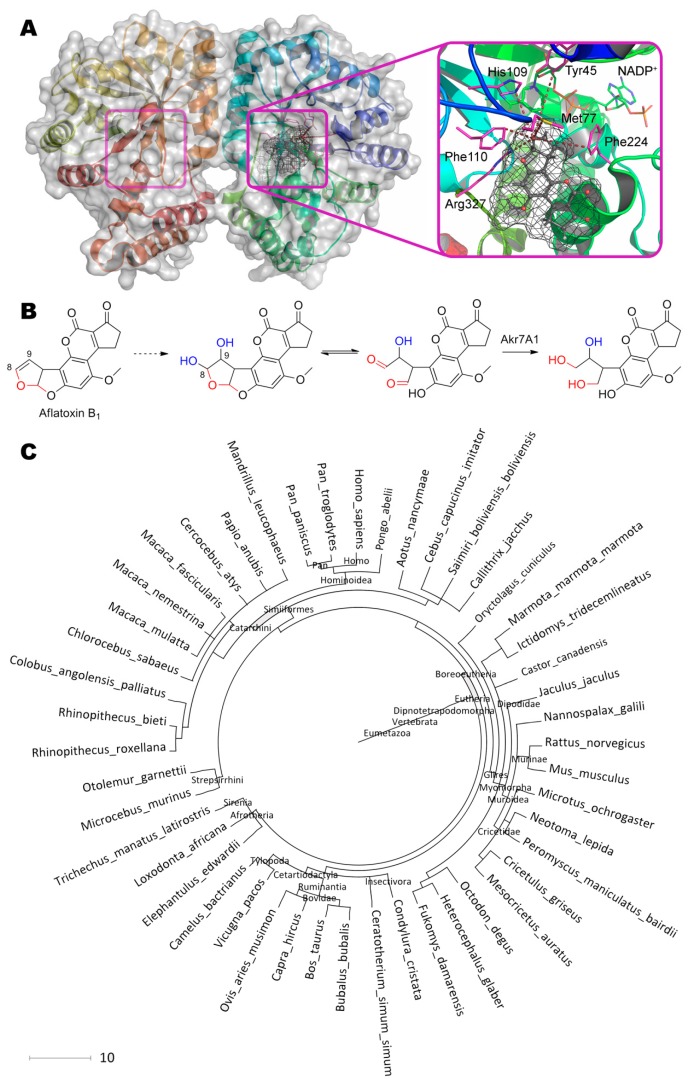
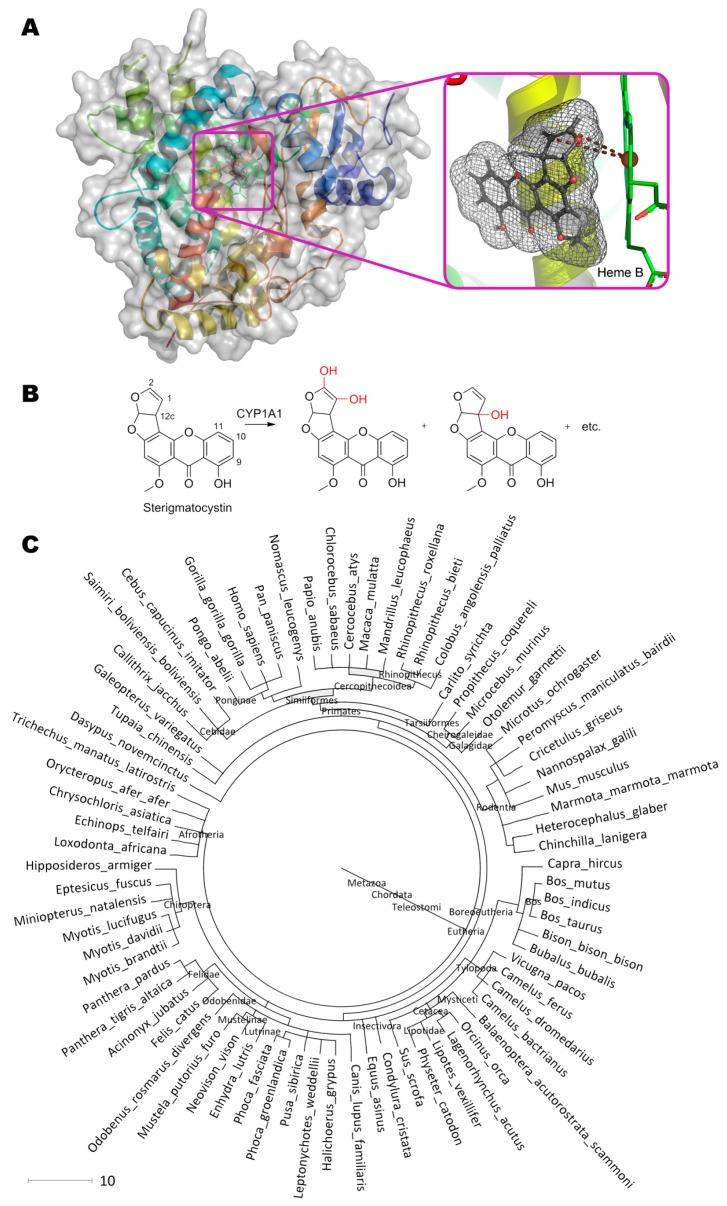
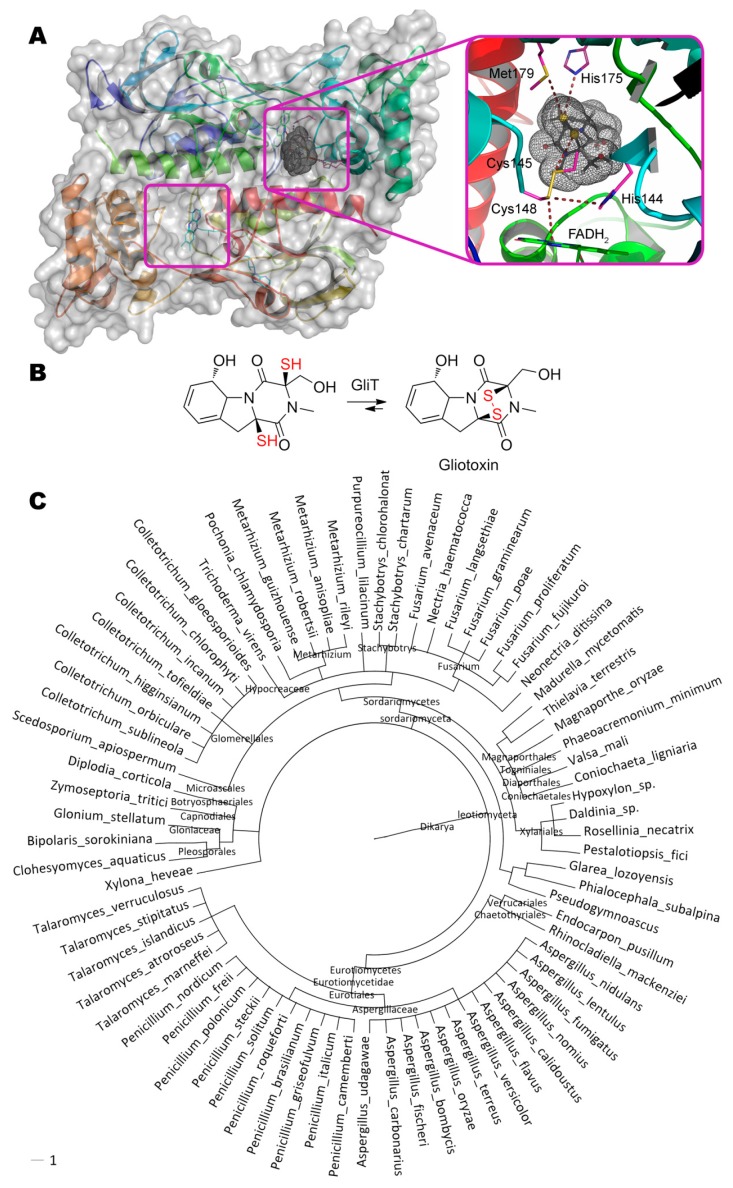
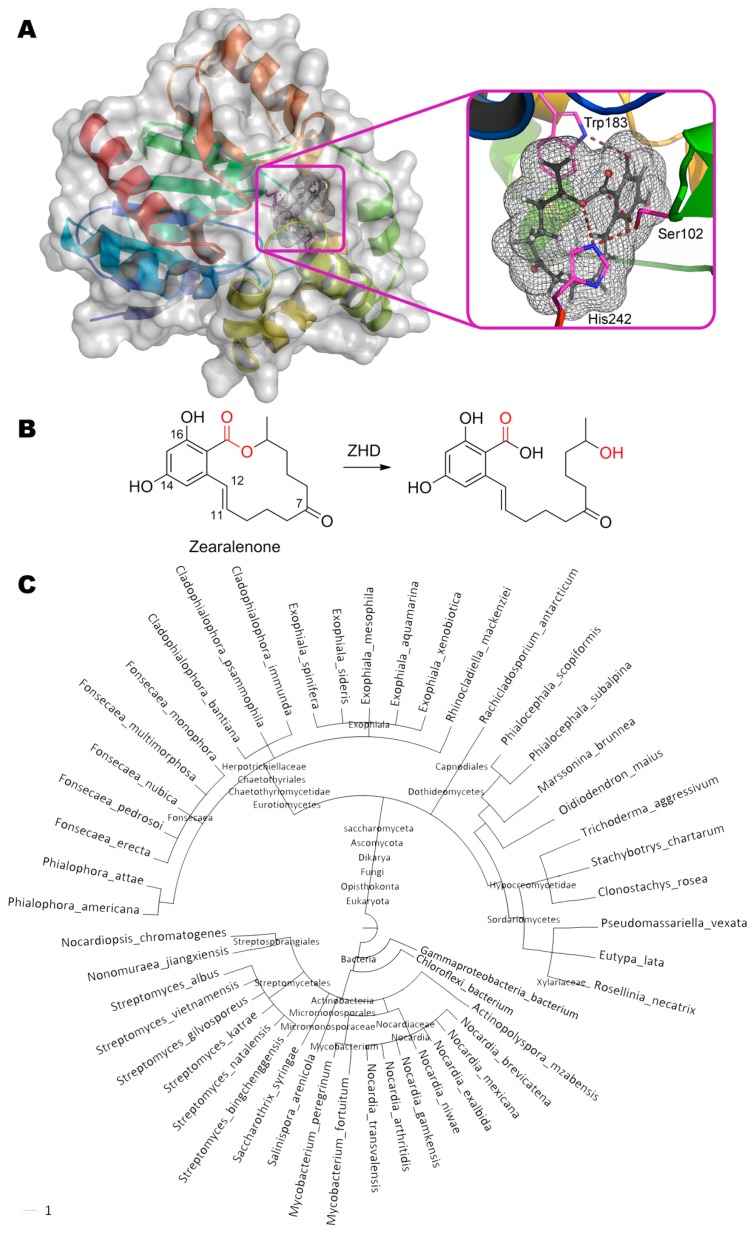
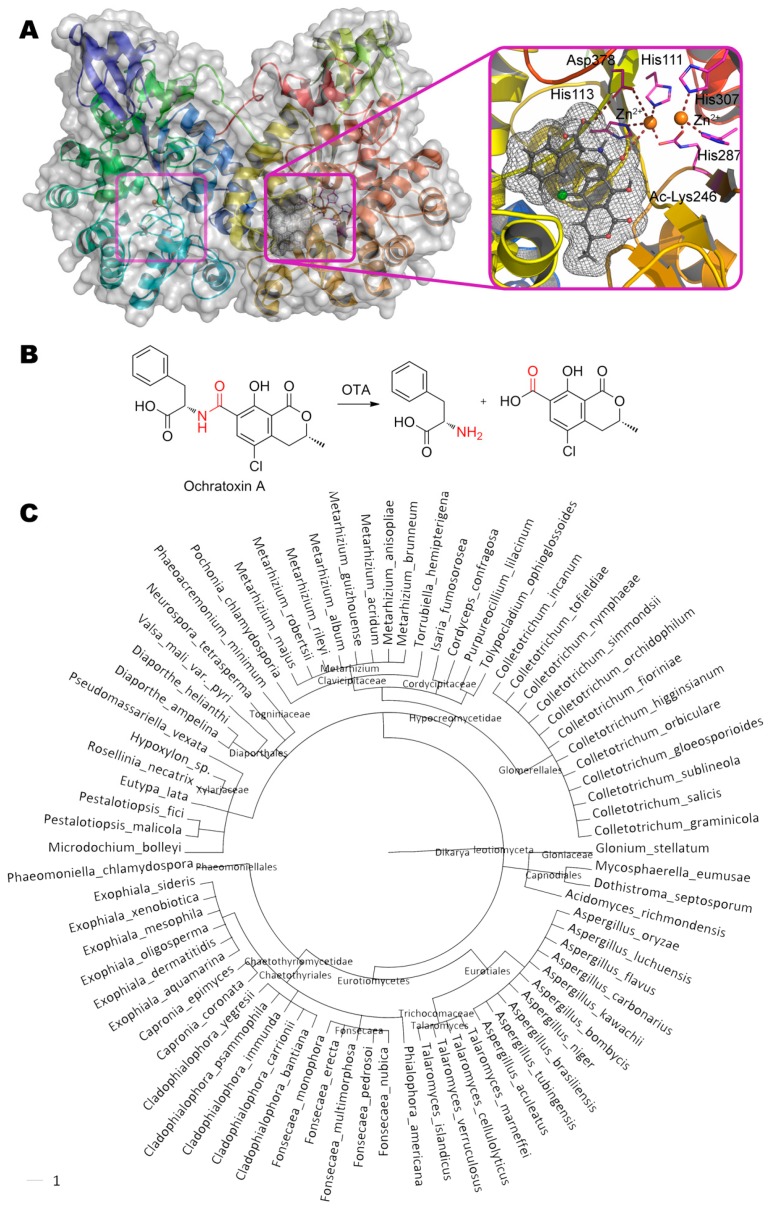
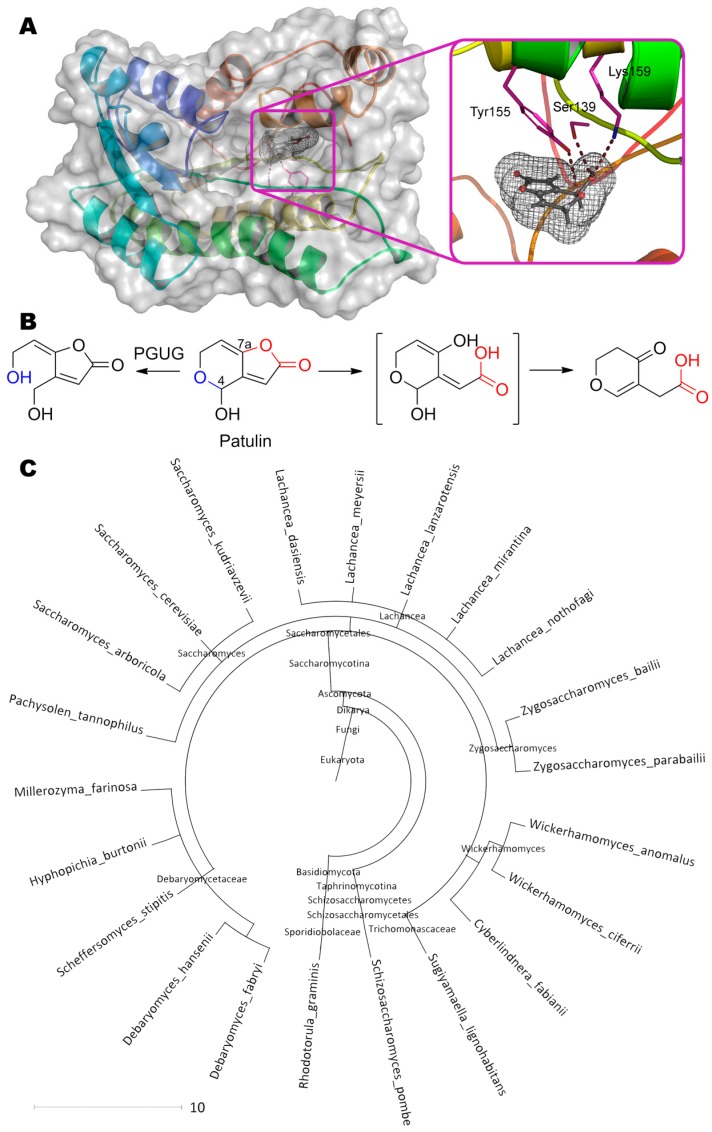
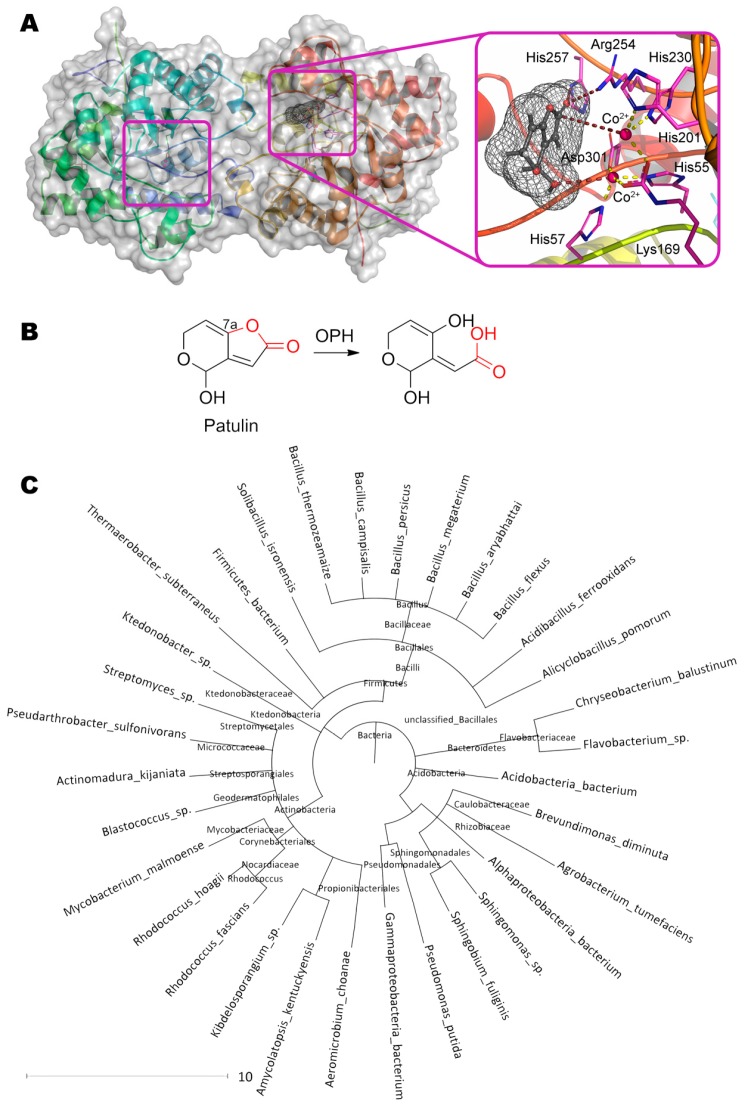
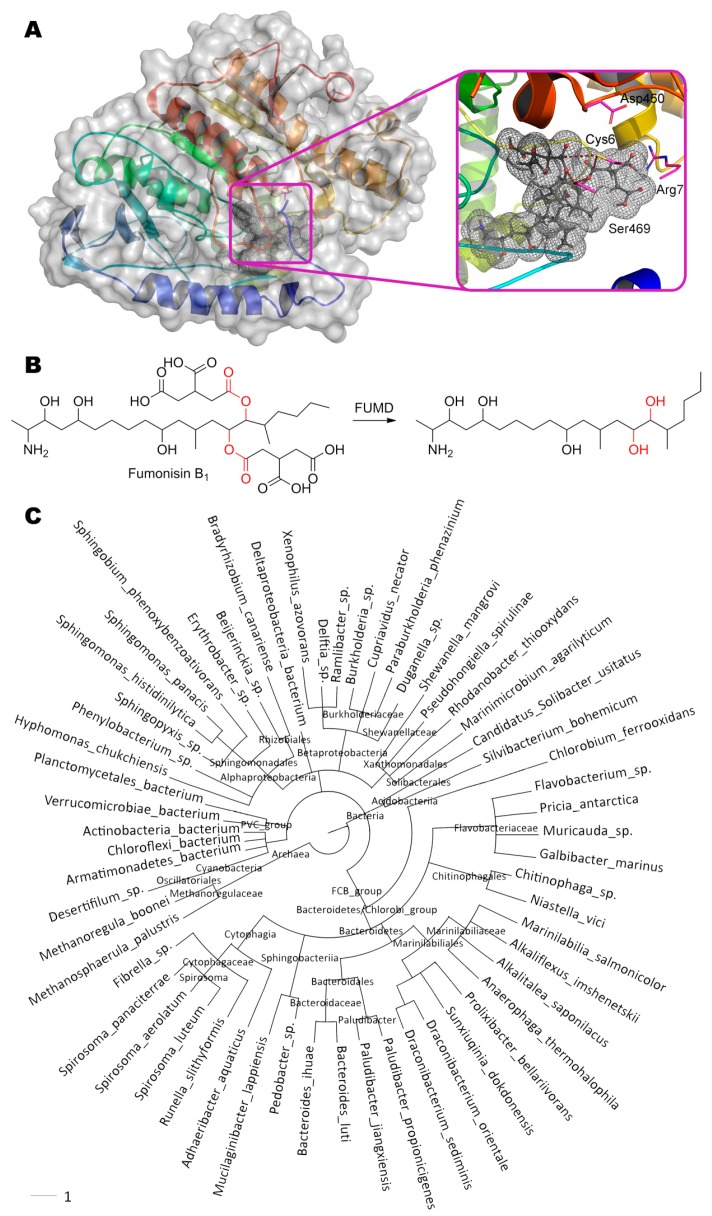
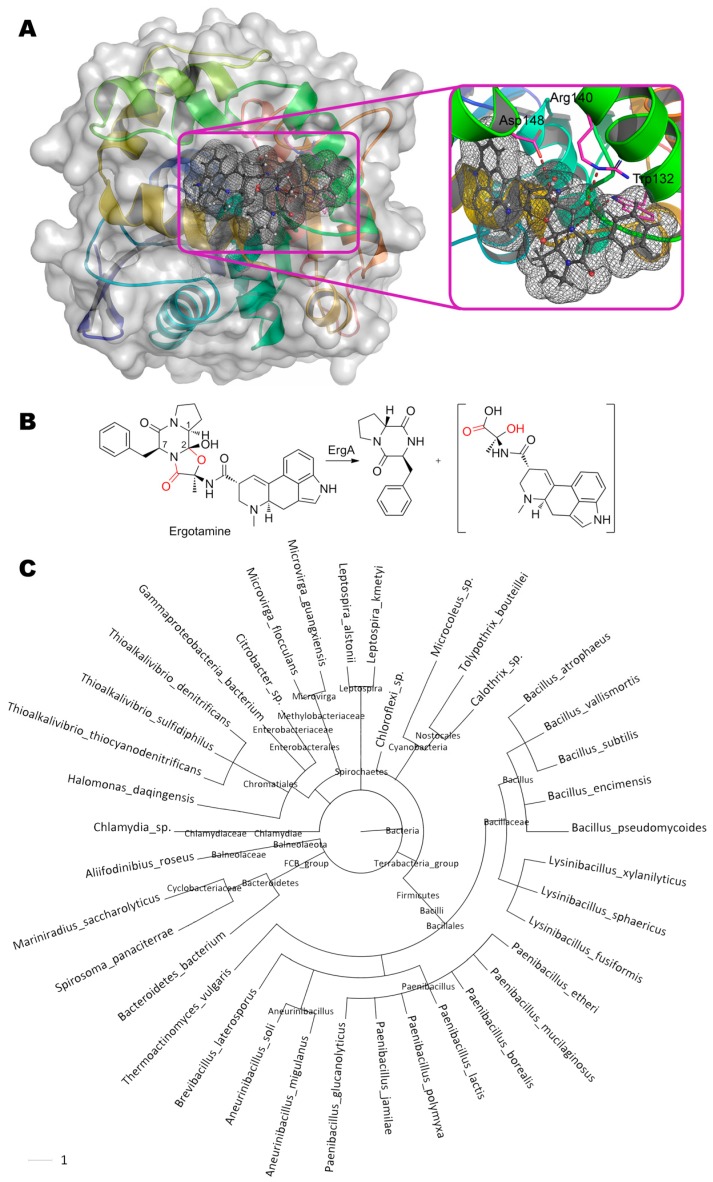

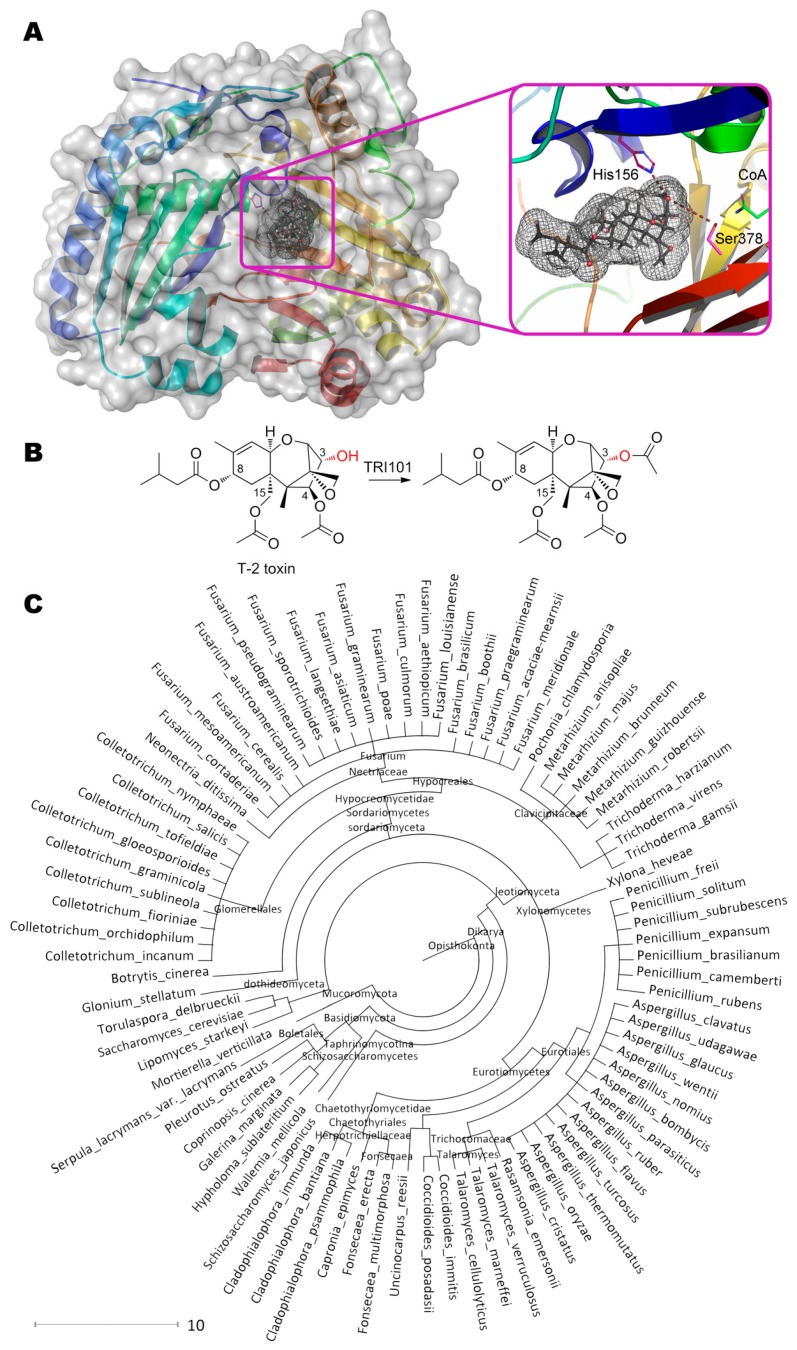
References
-
- Bryden W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012;173:134–158. doi: 10.1016/j.anifeedsci.2011.12.014. - DOI
-
- Temba B.A., Sultanbawa Y., Kriticos D.J., Fox G.P., Harvey J.J., Fletcher M.T. Tools for Defusing a Major Global Food and Feed Safety Risk: Nonbiological Postharvest Procedures to Decontaminate Mycotoxins in Foods and Feeds. J. Agric. Food Chem. 2016;64:8959–8972. doi: 10.1021/acs.jafc.6b03777. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
