

Deep Sequencing of Somatosensory Neurons Reveals Molecular Determinants of Intrinsic Physiological Properties
- PMID: 31248728
- PMCID: PMC6706313
- DOI: 10.1016/j.neuron.2019.05.039
Deep Sequencing of Somatosensory Neurons Reveals Molecular Determinants of Intrinsic Physiological Properties
Abstract
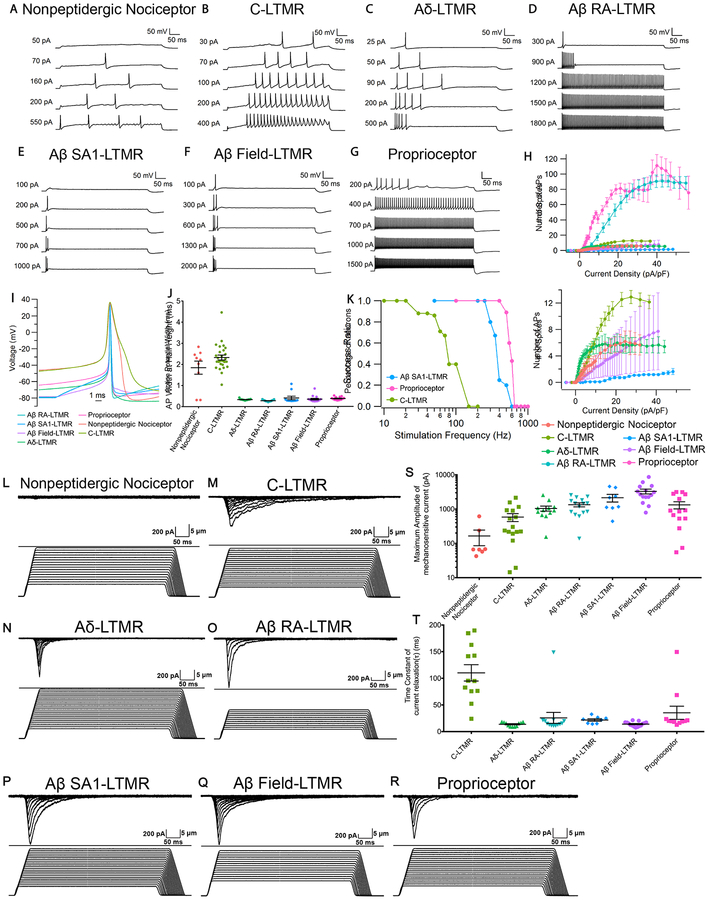
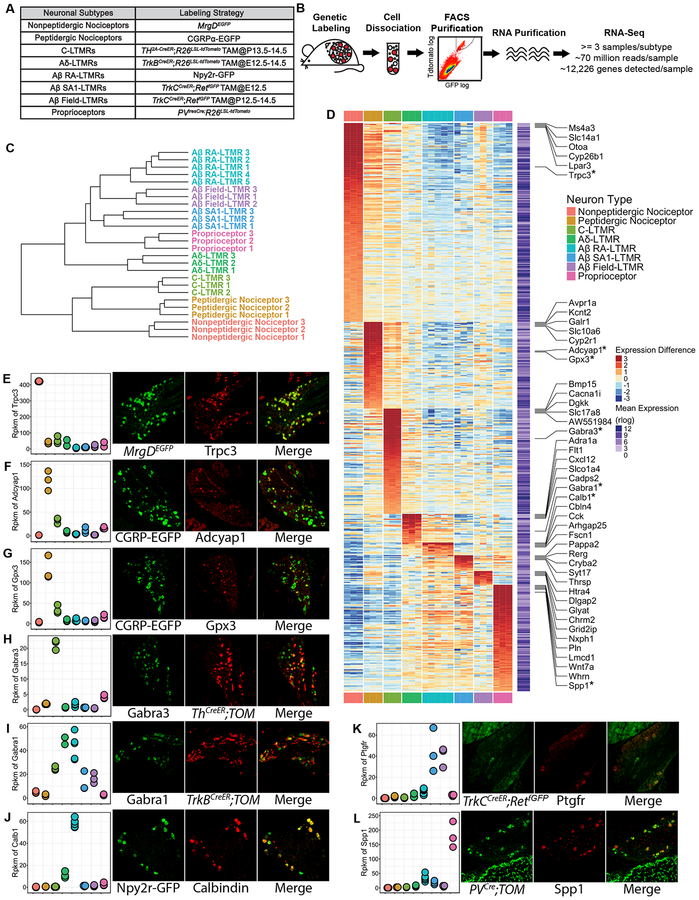
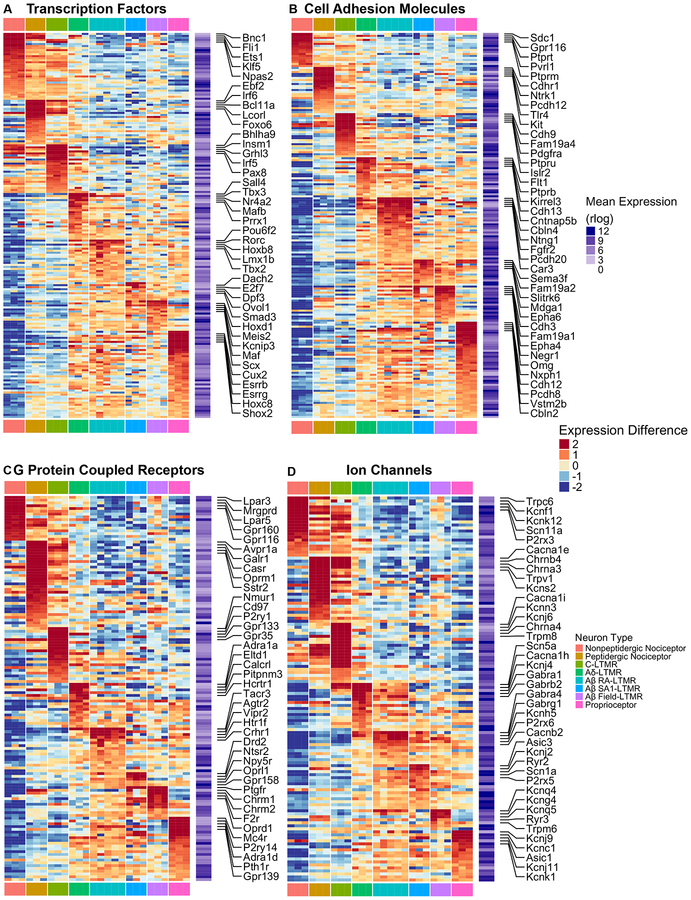
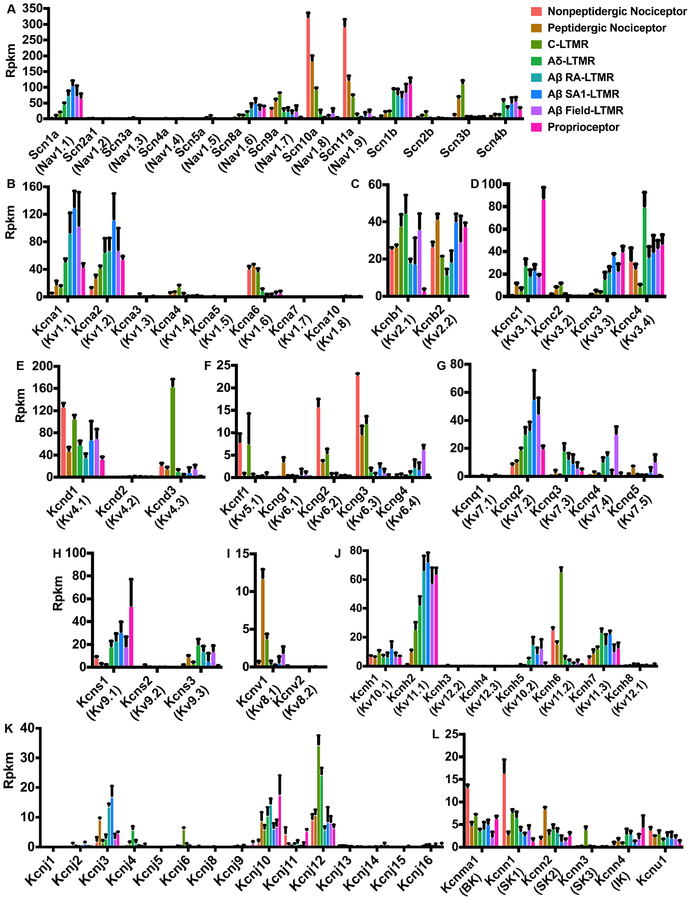
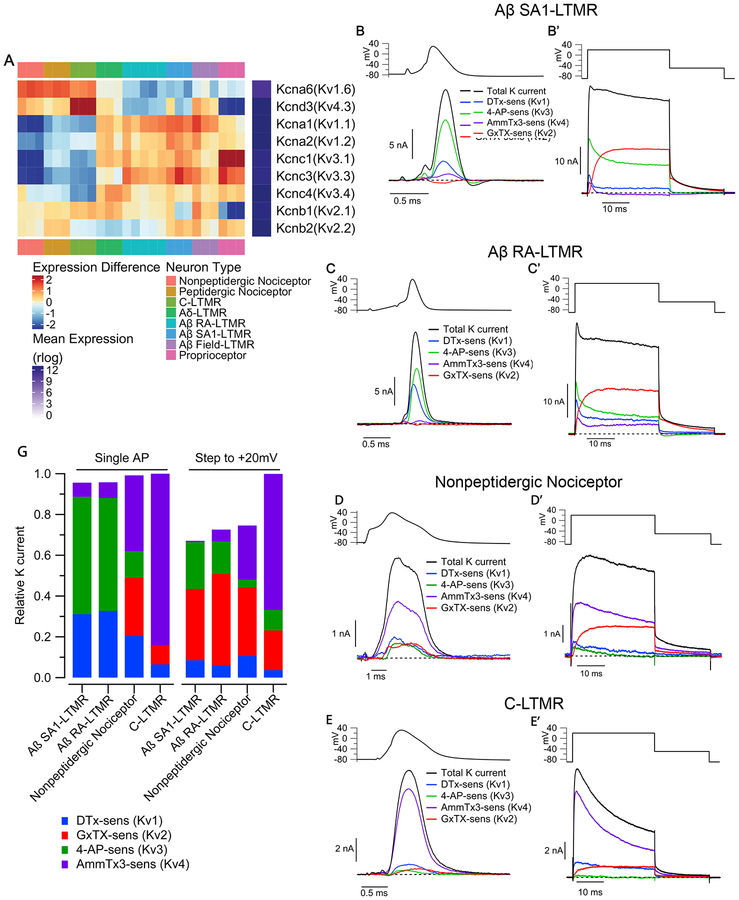
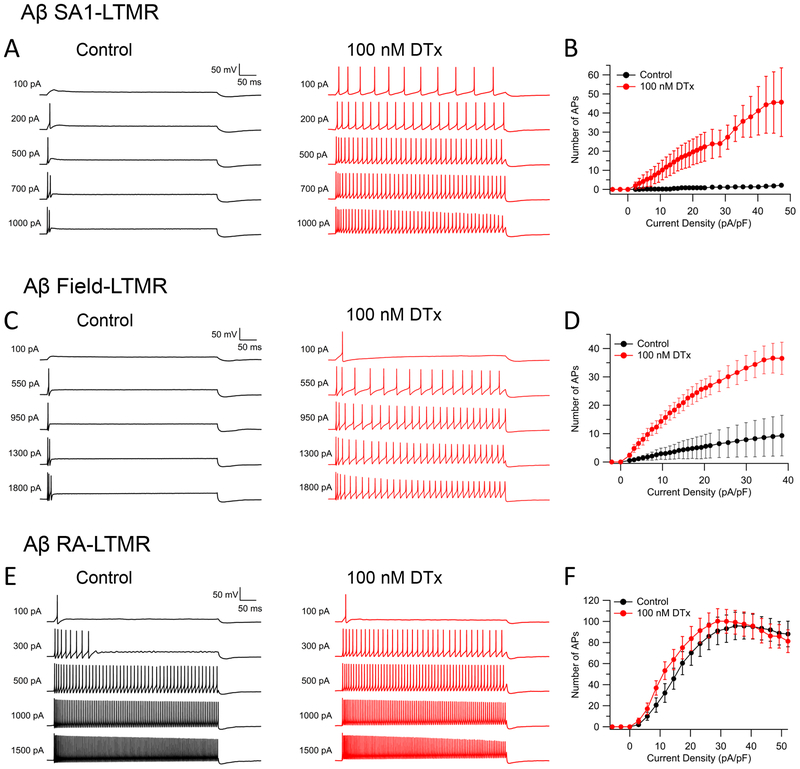
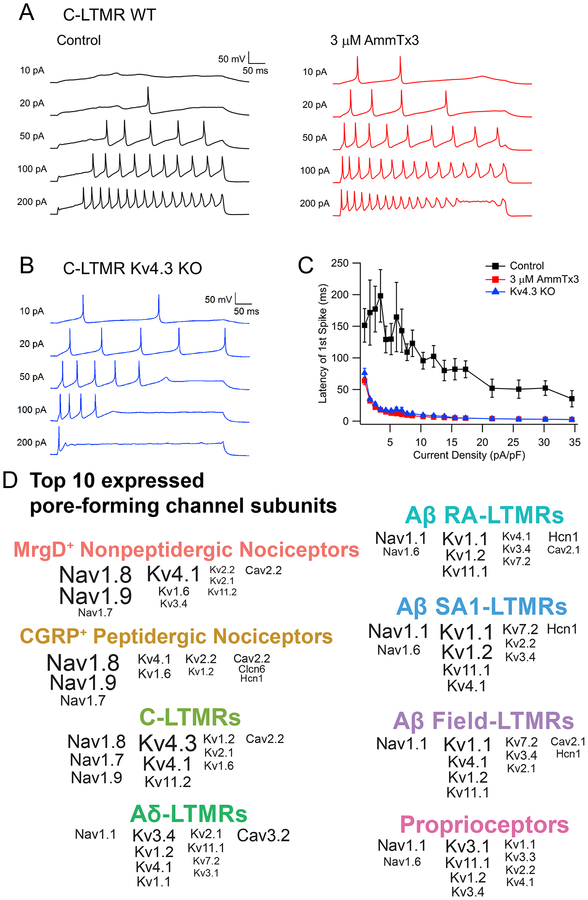
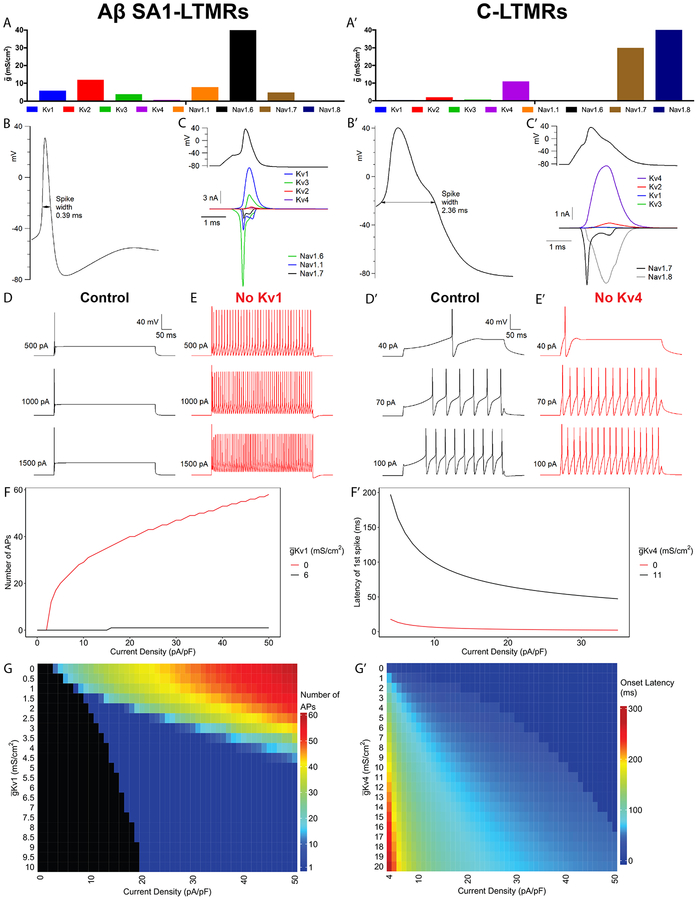
Dorsal root ganglion (DRG) sensory neuron subtypes defined by their in vivo properties display distinct intrinsic electrical properties. We used bulk RNA sequencing of genetically labeled neurons and electrophysiological analyses to define ion channel contributions to the intrinsic electrical properties of DRG neuron subtypes. The transcriptome profiles of eight DRG neuron subtypes revealed differentially expressed and functionally relevant genes, including voltage-gated ion channels. Guided by these data, electrophysiological analyses using pharmacological and genetic manipulations as well as computational modeling of DRG neuron subtypes were undertaken to assess the functions of select voltage-gated potassium channels (Kv1, Kv2, Kv3, and Kv4) in shaping action potential (AP) waveforms and firing patterns. Our findings show that the transcriptome profiles have predictive value for defining ion channel contributions to sensory neuron subtype-specific intrinsic physiological properties. The distinct ensembles of voltage-gated ion channels predicted to underlie the unique intrinsic physiological properties of eight DRG neuron subtypes are presented.
Keywords: DRG; RNA sequencing; genetic labeling; intrinsic membrane properties; mechanosensory neuron; somatosensation; transcriptome profile; voltage-gated ion channels.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interest
The authors declare no competing interests.
Figures
References
-
- Arcourt A, Gorham L, Dhandapani R, Prato V, Taberner FJ, Wende H, Gangadharan V, Birchmeier C, Heppenstall PA, and Lechner SG (2017). Touch Receptor-Derived Sensory Information Alleviates Acute Pain Signaling and Fine-Tunes Nociceptive Reflex Coordination. Neuron 93, 179–193. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
