

Substrate processing by the Cdc48 ATPase complex is initiated by ubiquitin unfolding
- PMID: 31249135
- PMCID: PMC6980381
- DOI: 10.1126/science.aax1033
Substrate processing by the Cdc48 ATPase complex is initiated by ubiquitin unfolding
Abstract
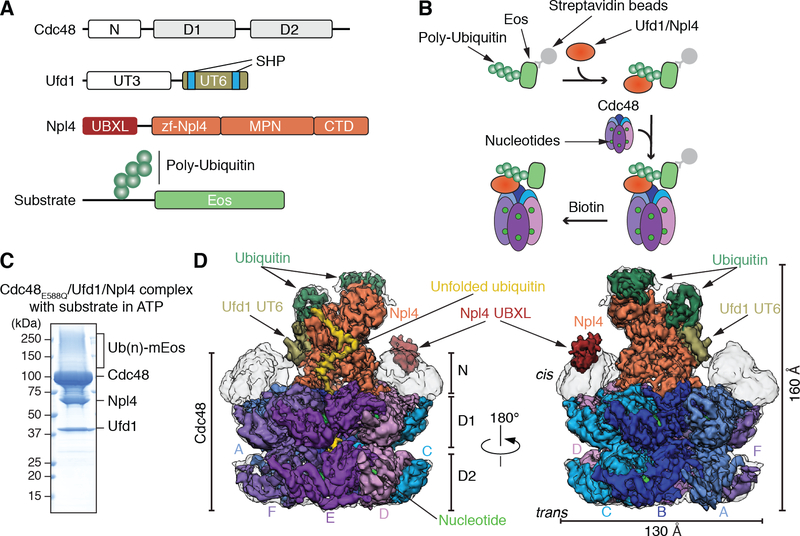
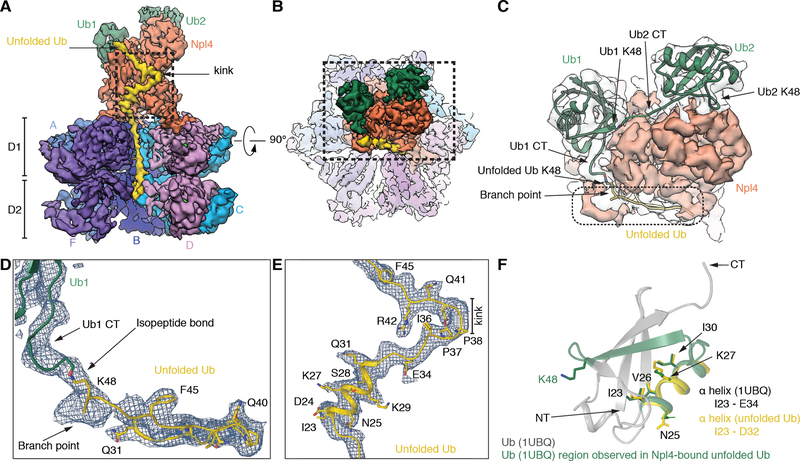
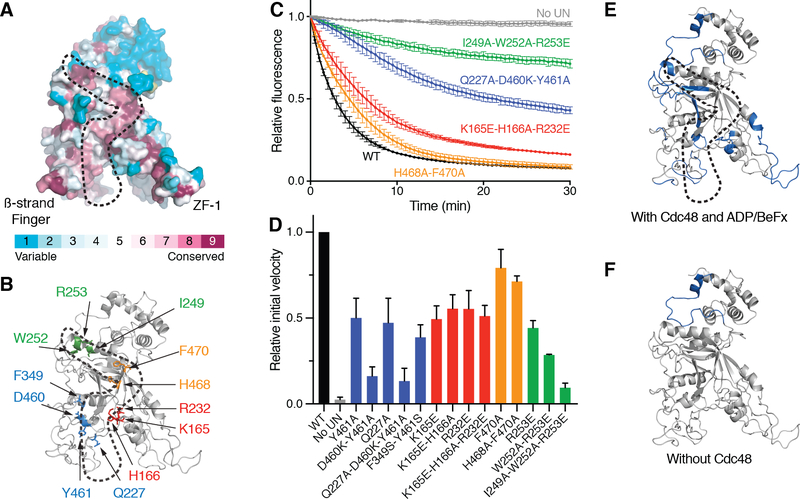
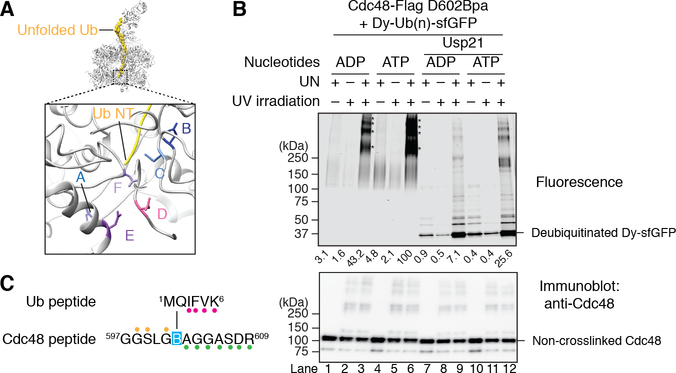
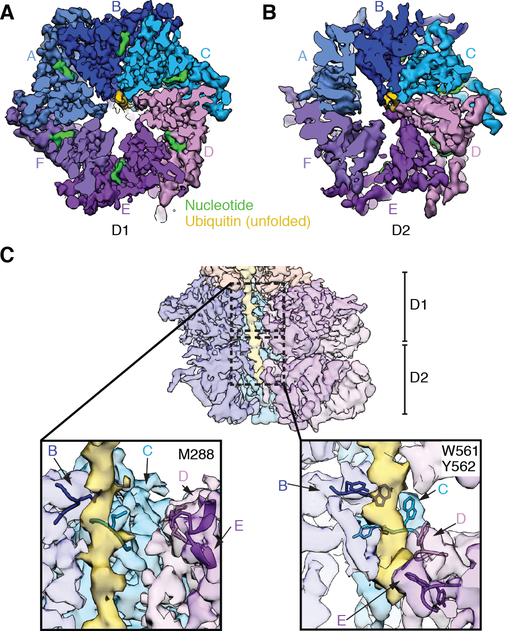
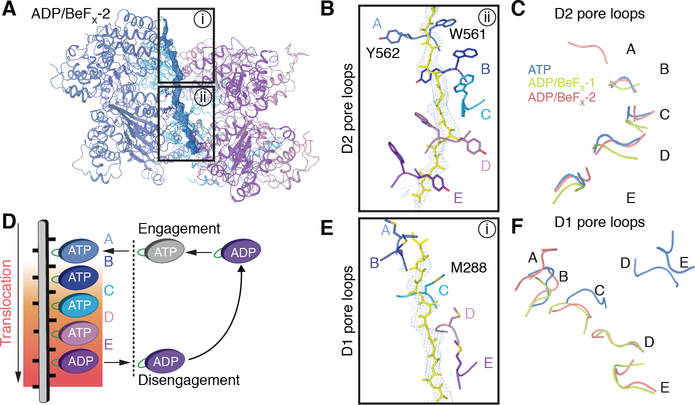
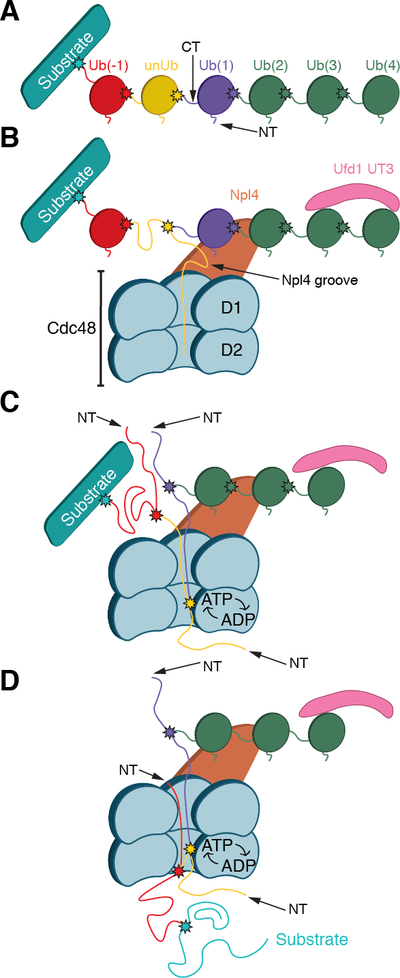
The Cdc48 adenosine triphosphatase (ATPase) (p97 or valosin-containing protein in mammals) and its cofactor Ufd1/Npl4 extract polyubiquitinated proteins from membranes or macromolecular complexes for subsequent degradation by the proteasome. How Cdc48 processes its diverse and often well-folded substrates is unclear. Here, we report cryo-electron microscopy structures of the Cdc48 ATPase in complex with Ufd1/Npl4 and polyubiquitinated substrate. The structures show that the Cdc48 complex initiates substrate processing by unfolding a ubiquitin molecule. The unfolded ubiquitin molecule binds to Npl4 and projects its N-terminal segment through both hexameric ATPase rings. Pore loops of the second ring form a staircase that acts as a conveyer belt to move the polypeptide through the central pore. Inducing the unfolding of ubiquitin allows the Cdc48 ATPase complex to process a broad range of substrates.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Similar articles
-
Translocation of polyubiquitinated protein substrates by the hexameric Cdc48 ATPase.Mol Cell. 2022 Feb 3;82(3):570-584.e8. doi: 10.1016/j.molcel.2021.11.033. Epub 2021 Dec 23. Mol Cell. 2022. PMID: 34951965 Free PMC article.
-
Structure of the Cdc48 ATPase with its ubiquitin-binding cofactor Ufd1-Npl4.Nat Struct Mol Biol. 2018 Jul;25(7):616-622. doi: 10.1038/s41594-018-0085-x. Epub 2018 Jul 2. Nat Struct Mol Biol. 2018. PMID: 29967539 Free PMC article.
-
The Ufd1 cofactor determines the linkage specificity of polyubiquitin chain engagement by the AAA+ ATPase Cdc48.Mol Cell. 2023 Mar 2;83(5):759-769.e7. doi: 10.1016/j.molcel.2023.01.016. Epub 2023 Feb 2. Mol Cell. 2023. PMID: 36736315 Free PMC article.
-
Cdc48-Ufd1-Npl4: stuck in the middle with Ub.Curr Biol. 2002 May 14;12(10):R366-71. doi: 10.1016/s0960-9822(02)00862-x. Curr Biol. 2002. PMID: 12015140 Review.
-
Toward an understanding of the Cdc48/p97 ATPase.F1000Res. 2017 Aug 3;6:1318. doi: 10.12688/f1000research.11683.1. eCollection 2017. F1000Res. 2017. PMID: 28815021 Free PMC article. Review.
Cited by
-
Substrate recognition mechanism of the endoplasmic reticulum-associated ubiquitin ligase Doa10.bioRxiv [Preprint]. 2024 Jan 9:2024.01.09.574907. doi: 10.1101/2024.01.09.574907. bioRxiv. 2024. Update in: Nat Commun. 2024 Mar 11;15(1):2182. doi: 10.1038/s41467-024-46409-2. PMID: 38260251 Free PMC article. Updated. Preprint.
-
Valosin-Containing Protein (VCP)/p97 Oligomerization.Subcell Biochem. 2024;104:485-501. doi: 10.1007/978-3-031-58843-3_18. Subcell Biochem. 2024. PMID: 38963497 Review.
-
Stairway to translocation: AAA+ motor structures reveal the mechanisms of ATP-dependent substrate translocation.Protein Sci. 2020 Feb;29(2):407-419. doi: 10.1002/pro.3743. Epub 2019 Oct 17. Protein Sci. 2020. PMID: 31599052 Free PMC article. Review.
-
USP7 and VCPFAF1 define the SUMO/Ubiquitin landscape at the DNA replication fork.Cell Rep. 2021 Oct 12;37(2):109819. doi: 10.1016/j.celrep.2021.109819. Cell Rep. 2021. PMID: 34644576 Free PMC article.
-
Structural and Functional Analysis of Disease-Linked p97 ATPase Mutant Complexes.Int J Mol Sci. 2021 Jul 28;22(15):8079. doi: 10.3390/ijms22158079. Int J Mol Sci. 2021. PMID: 34360842 Free PMC article.
References
-
- van den Boom J, Meyer H, VCP/p97-Mediated Unfolding as a Principle in Protein Homeostasis and Signaling. Mol. Cell 69, 182–194 (2018). - PubMed
-
- Watts GDJ et al., Inclusion body myopathy associated with Paget disease of bone and frontotemporal dementia is caused by mutant valosin-containing protein. Nat. Genet 36, 377–381 (2004). - PubMed
-
- Kimonis VE, Fulchiero E, Vesa J, Watts G, VCP disease associated with myopathy, Paget disease of bone and frontotemporal dementia: Review of a unique disorder. Biochim. Biophys. Acta - Mol. Basis Dis 1782, 744–748 (2008). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
