

Identification of salicylic acid-independent responses in an Arabidopsis phosphatidylinositol 4-kinase beta double mutant
- PMID: 31250883
- PMCID: PMC7182594
- DOI: 10.1093/aob/mcz112
Identification of salicylic acid-independent responses in an Arabidopsis phosphatidylinositol 4-kinase beta double mutant
Abstract
Background and aims: We have recently shown that an Arabidopsis thaliana double mutant of type III phosphatidylinositol-4-kinases (PI4Ks), pi4kβ1β2, constitutively accumulated a high level of salicylic acid (SA). By crossing this pi4kβ1β2 double mutant with mutants impaired in SA synthesis (such as sid2 impaired in isochorismate synthase) or transduction, we demonstrated that the high SA level was responsible for the dwarfism phenotype of the double mutant. Here we aimed to distinguish between the SA-dependent and SA-independent effects triggered by the deficiency in PI4Kβ1 and PI4Kβ2.
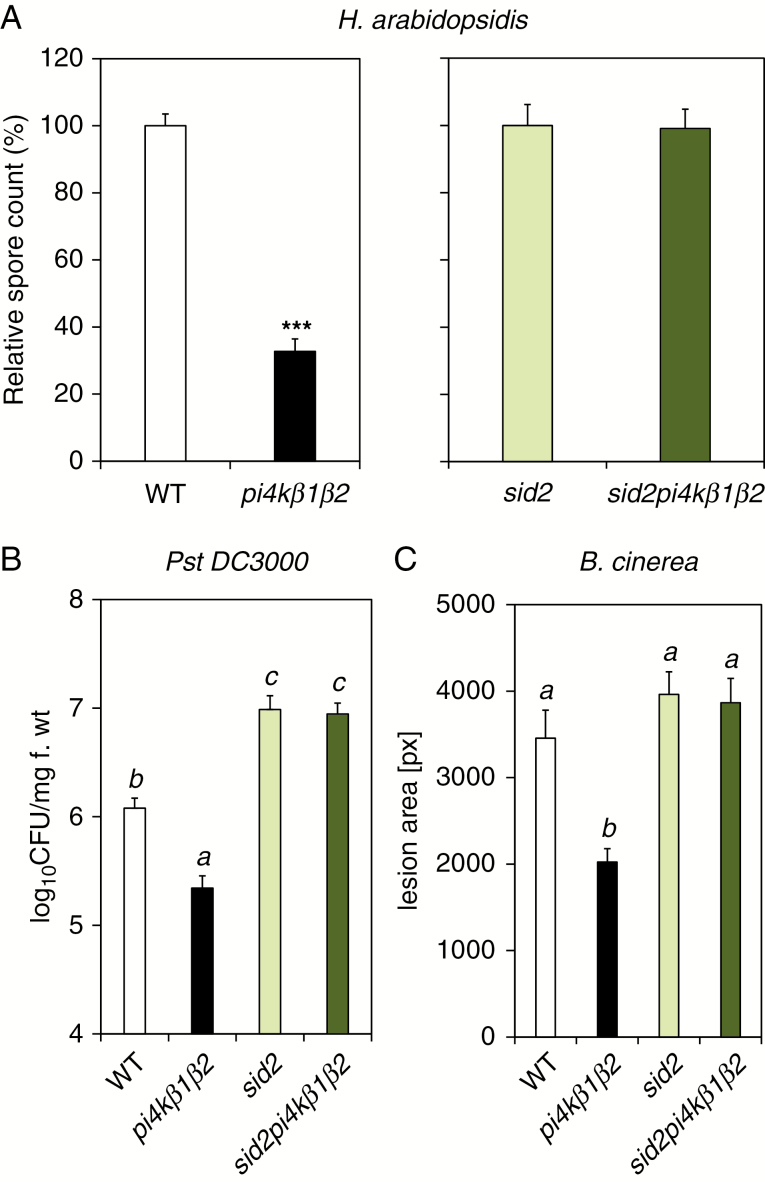
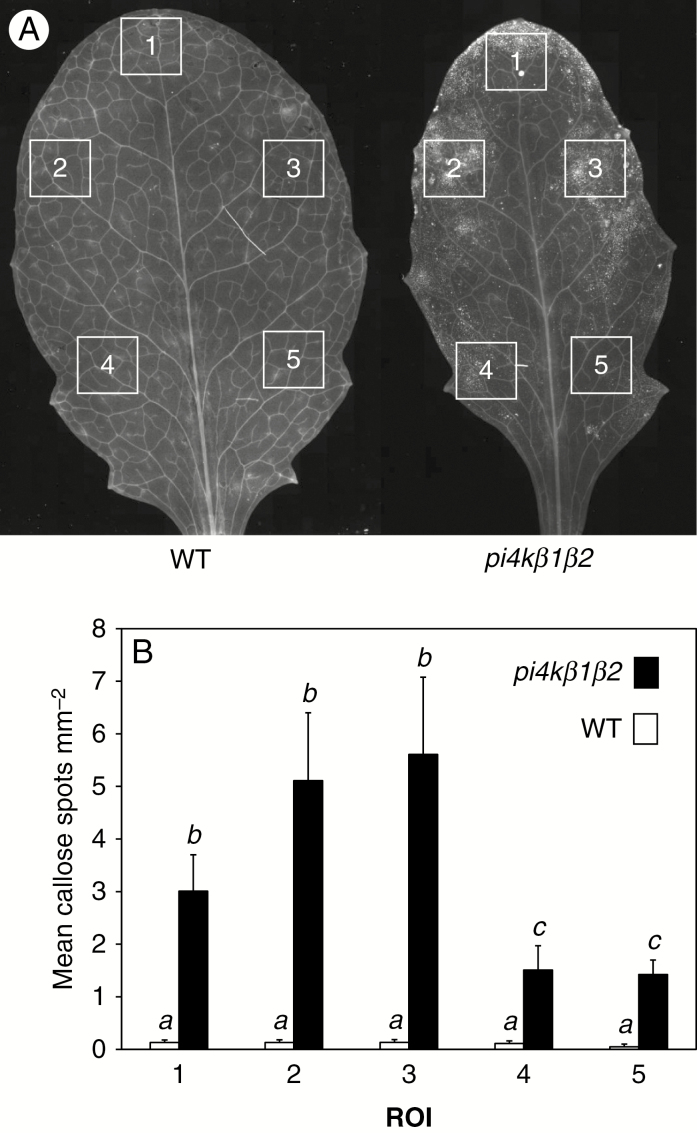
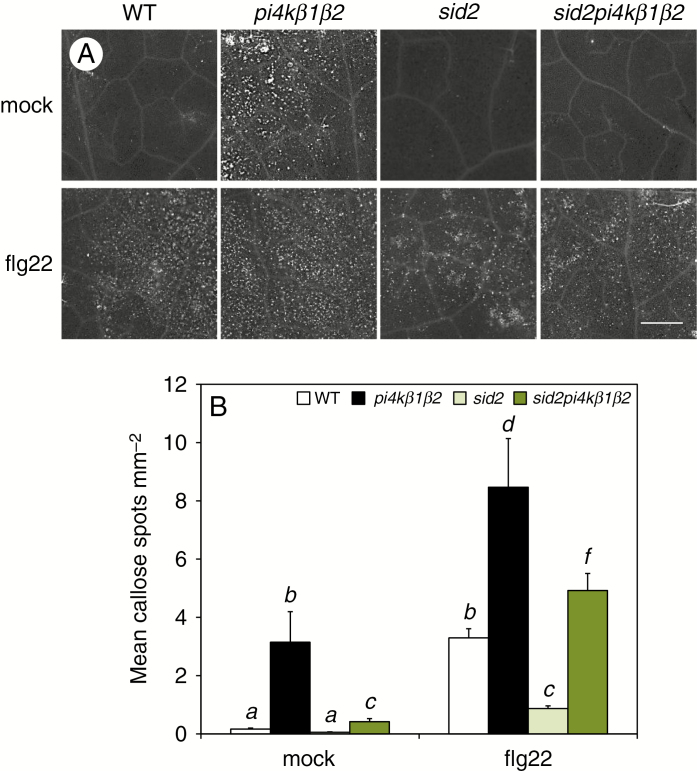
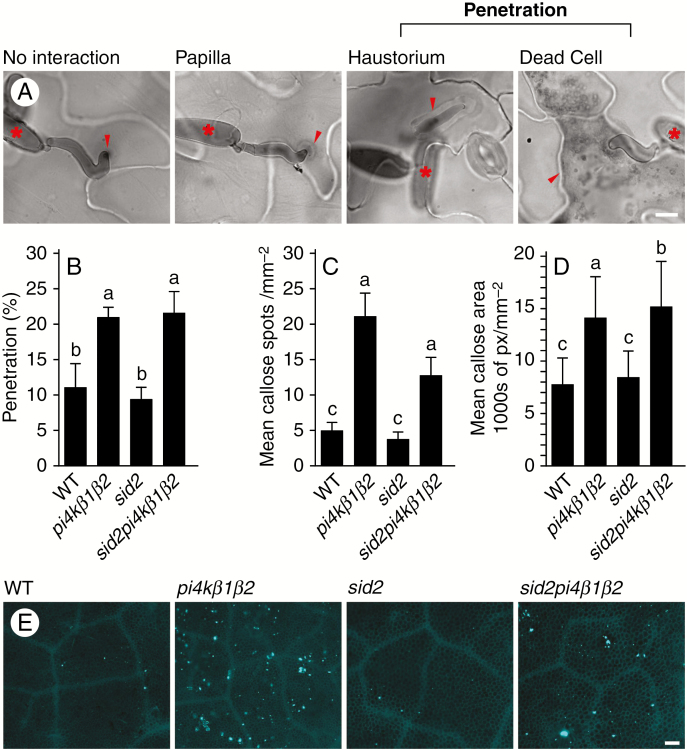
Methods: To achieve this we used the sid2pi4kβ1β2 triple mutant. High-throughput analyses of phytohormones were performed on this mutant together with pi4kβ1β2 and sid2 mutants and wild-type plants. Responses to pathogens, namely Hyaloperonospora arabidopsidis, Pseudomonas syringae and Botrytis cinerea, and also to the non-host fungus Blumeria graminis, were also determined. Callose accumulation was monitored in response to flagellin.
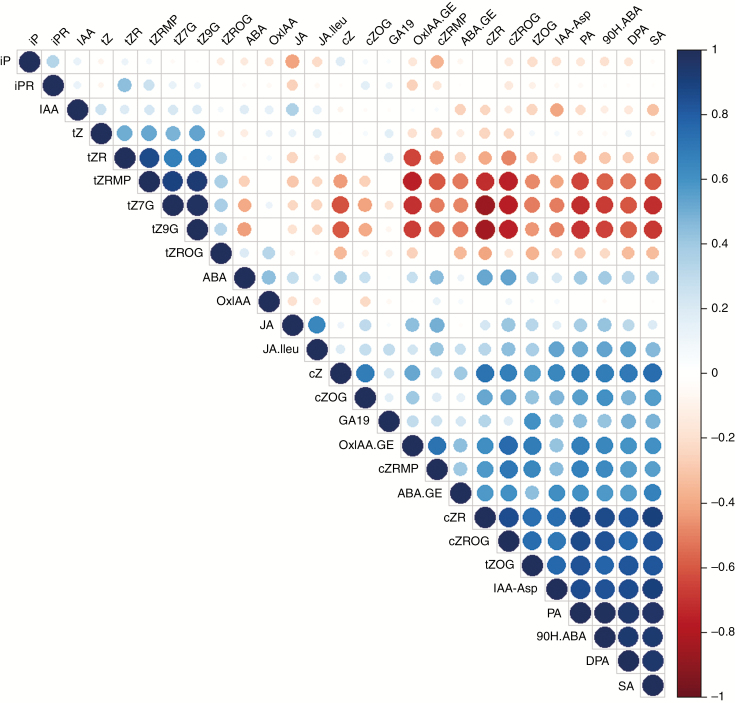
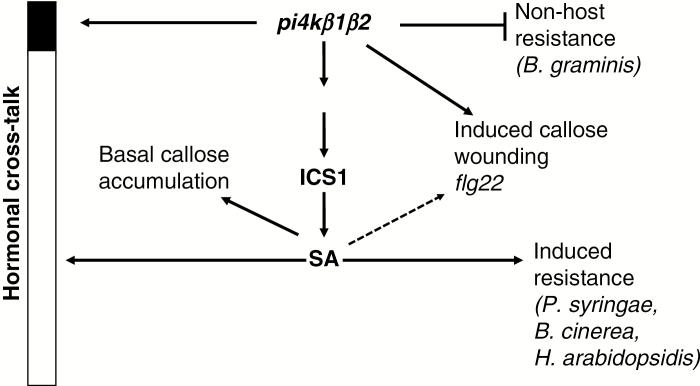
Key results: We show here the prominent role of high SA levels in influencing the concentration of many other tested phytohormones, including abscisic acid and its derivatives, the aspartate-conjugated form of indole-3-acetic acid and some cytokinins such as cis-zeatin. We show that the increased resistance of pi4kβ1β2 plants to the host pathogens H. arabidopsidis, P. syringae pv. tomato DC3000 and Bothrytis cinerea is dependent on accumulation of high SA levels. In contrast, accumulation of callose in pi4kβ1β2 after flagellin treatment was independent of SA. Concerning the response to Blumeria graminis, both callose accumulation and fungal penetration were enhanced in the pi4kβ1β2 double mutant compared to wild-type plants. Both of these processes occurred in an SA-independent manner.
Conclusions: Our data extensively illustrate the influence of SA on other phytohormone levels. The sid2pi4kβ1β2 triple mutant revealed the role of PI4Kβ1/β2 per se, thus showing the importance of these enzymes in plant defence responses.
Keywords: Arabidopsis thaliana; biotic stress; callose; isochorismate synthase 1; pathogens; phytohormones; pi4kβ1β2/PI4Ks; salicylic acid.
© The Author(s) 2019. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
References
-
- Antignani V, Klocko AL, Bak G, Chandrasekaran SD, Dunivin T, Nielsen E. 2015. Recruitment of PLANT U-BOX13 and the PI4Kbeta1/beta2 phosphatidylinositol-4 kinases by the small GTPase RabA4B plays important roles during salicylic acid-mediated plant defense signaling in Arabidopsis. Plant Cell 27: 243–261. - PMC - PubMed
-
- Cao H, Glazebrook J, Clarke JD, Volko S, Dong X. 1997. The Arabidopsis NPR1 gene that controls systemic acquired resistance encodes a novel protein containing ankyrin repeats. Cell 88: 57–63. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
