

Interactions between a subset of substrate side chains and AAA+ motor pore loops determine grip during protein unfolding
- PMID: 31251172
- PMCID: PMC6677533
- DOI: 10.7554/eLife.46808
Interactions between a subset of substrate side chains and AAA+ motor pore loops determine grip during protein unfolding
Abstract
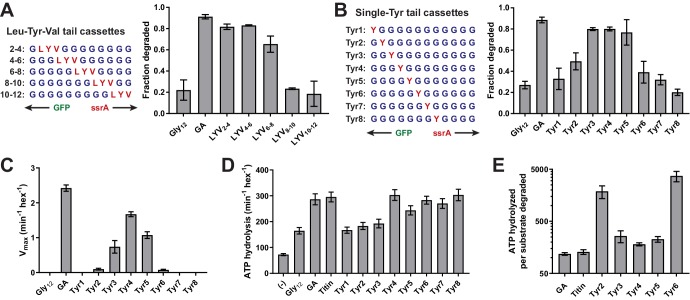
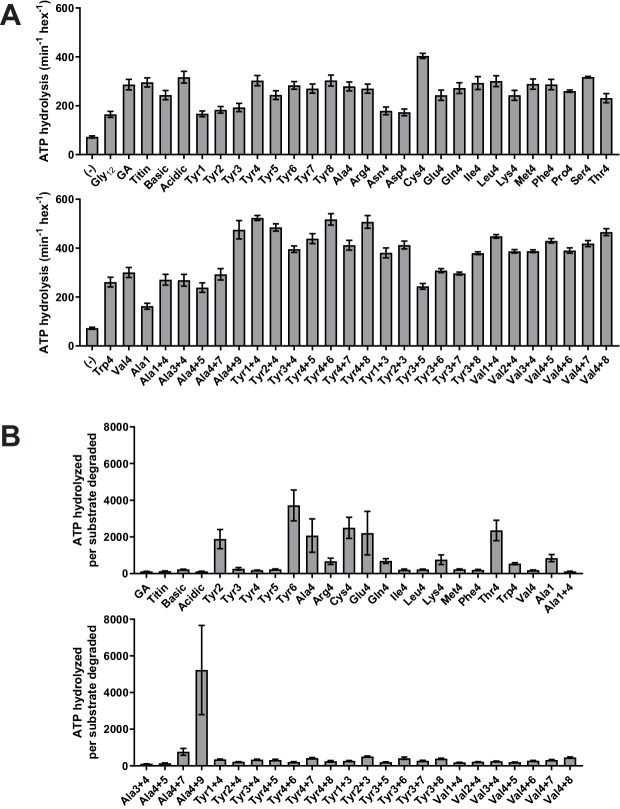
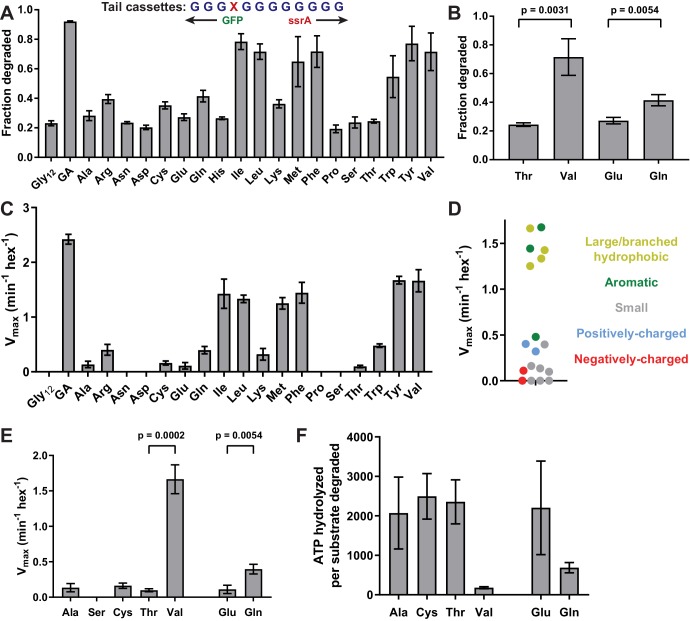
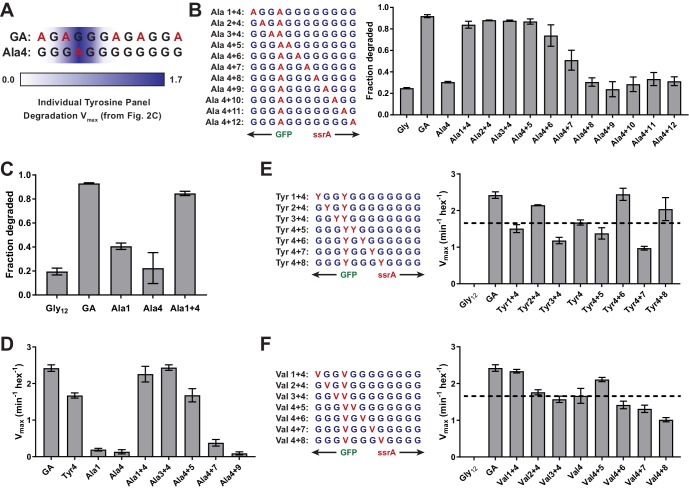
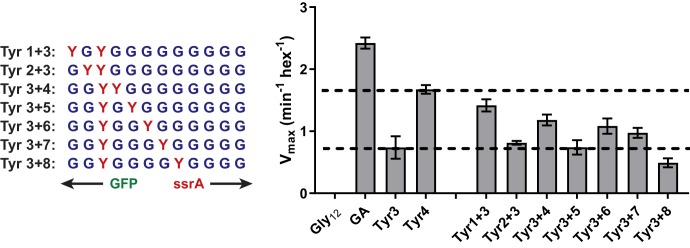
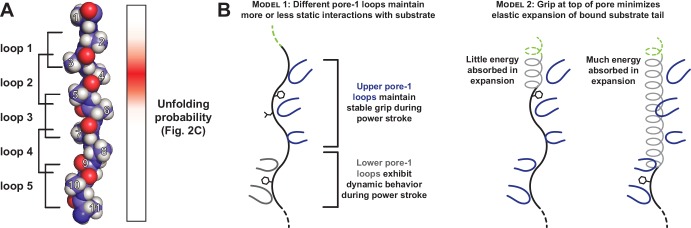
Most AAA+ remodeling motors denature proteins by pulling on the peptide termini of folded substrates, but it is not well-understood how motors produce grip when resisting a folded domain. Here, at single amino-acid resolution, we identify the determinants of grip by measuring how substrate tail sequences alter the unfolding activity of the unfoldase-protease ClpXP. The seven amino acids abutting a stable substrate domain are key, with residues 2-6 forming a core that contributes most significantly to grip. ClpX grips large hydrophobic and aromatic side chains strongly and small, polar, or charged side chains weakly. Multiple side chains interact with pore loops synergistically to strengthen grip. In combination with recent structures, our results support a mechanism in which unfolding grip is primarily mediated by non-specific van der Waal's interactions between core side chains of the substrate tail and a subset of YVG loops at the top of the ClpX axial pore.
Keywords: AAA+ enzymes; ATP-dependent protein unfolding; ClpXP; E. coli; biochemistry; chemical biology; molecular biophysics; protein degradation; structural biology.
© 2019, Bell et al.
Conflict of interest statement
TB, TB, RS No competing interests declared
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
