

Mechanisms of PINK1, ubiquitin and Parkin interactions in mitochondrial quality control and beyond
- PMID: 31254044
- PMCID: PMC11105328
- DOI: 10.1007/s00018-019-03203-4
Mechanisms of PINK1, ubiquitin and Parkin interactions in mitochondrial quality control and beyond
Abstract
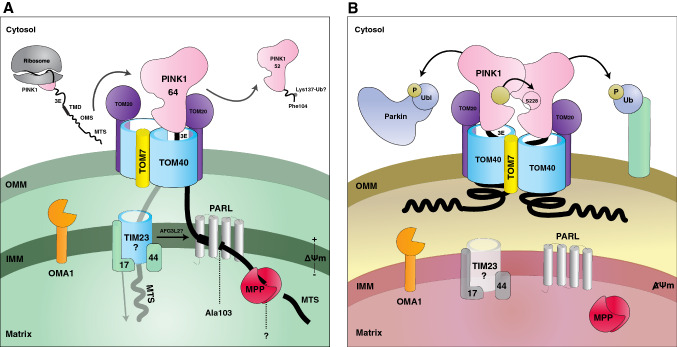
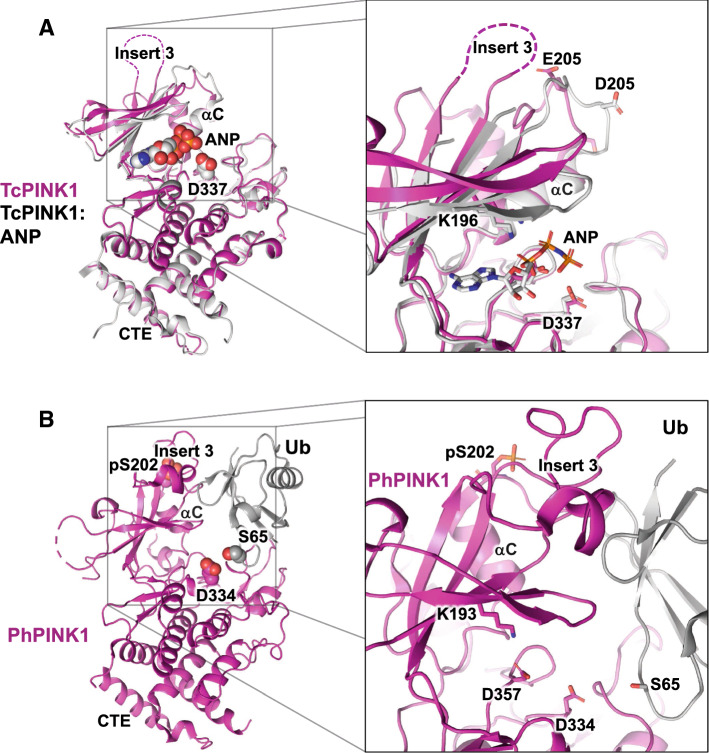
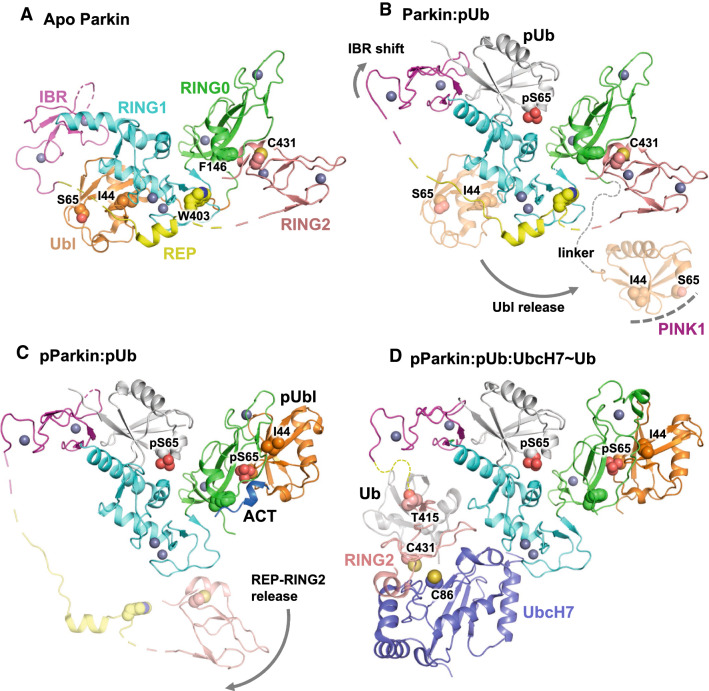
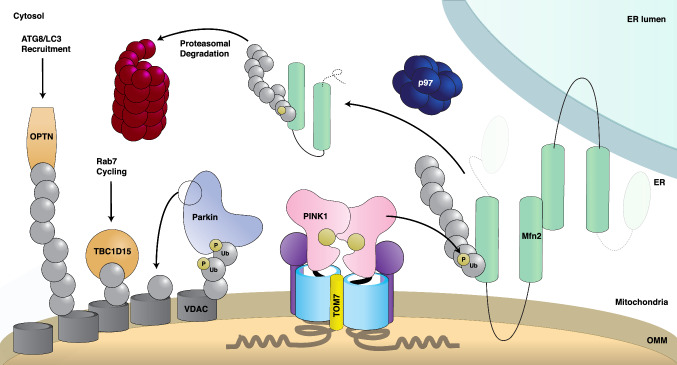
Parkinson's disease (PD) is a degenerative movement disorder resulting from the loss of specific neuron types in the midbrain. Early environmental and pathophysiological studies implicated mitochondrial damage and protein aggregation as the main causes of PD. These findings are now vindicated by the characterization of more than 20 genes implicated in rare familial forms of the disease. In particular, two proteins encoded by the Parkin and PINK1 genes, whose mutations cause early-onset autosomal recessive PD, function together in a mitochondrial quality control pathway. In this review, we will describe recent development in our understanding of their mechanisms of action, structure, and function. We explain how PINK1 acts as a mitochondrial damage sensor via the regulated proteolysis of its N-terminus and the phosphorylation of ubiquitin tethered to outer mitochondrial membrane proteins. In turn, phospho-ubiquitin recruits and activates Parkin via conformational changes that increase its ubiquitin ligase activity. We then describe how the formation of polyubiquitin chains on mitochondria triggers the recruitment of the autophagy machinery or the formation of mitochondria-derived vesicles. Finally, we discuss the evidence for the involvement of these mechanisms in physiological processes such as immunity and inflammation, as well as the links to other PD genes.
Keywords: Kinase; Mitochondria; PINK1; Parkin; Parkinson; Ubiquitin.
Conflict of interest statement
ANB declares no conflicts of interest. J-FT is a consultant for Mitokinin Inc. and a founding member of M4ND Pharma Inc.
Figures
References
-
- Spillantini MG, Schmidt ML, Lee VM, Trojanowski JQ, Jakes R, Goedert M. Alpha-synuclein in Lewy bodies. Nature. 1997;388:839–840. - PubMed
-
- Langston JW, Ballard PA., Jr Parkinson’s disease in a chemist working with 1-methyl-4-phenyl-1,2,5,6-tetrahydropyridine. N Engl J Med. 1983;309:310. - PubMed
-
- Langston JW, Ballard P, Tetrud JW, Irwin I. Chronic Parkinsonism in humans due to a product of meperidine-analog synthesis. Science. 1983;219:979–980. - PubMed
-
- Schapira AH, Cooper JM, Dexter D, Jenner P, Clark JB, Marsden CD. Mitochondrial complex I deficiency in Parkinson’s disease. Lancet. 1989;1:1269. - PubMed
-
- Bender A, Krishnan KJ, Morris CM, Taylor GA, Reeve AK, Perry RH, Jaros E, Hersheson JS, Betts J, Klopstock T, Taylor RW, Turnbull DM. High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease. Nat Genet. 2006;38:515–517. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
