

Evidence for bidirectional and trans-synaptic parasympathetic and sympathetic propagation of alpha-synuclein in rats
- PMID: 31254094
- PMCID: PMC6778265
- DOI: 10.1007/s00401-019-02040-w
Evidence for bidirectional and trans-synaptic parasympathetic and sympathetic propagation of alpha-synuclein in rats
Abstract
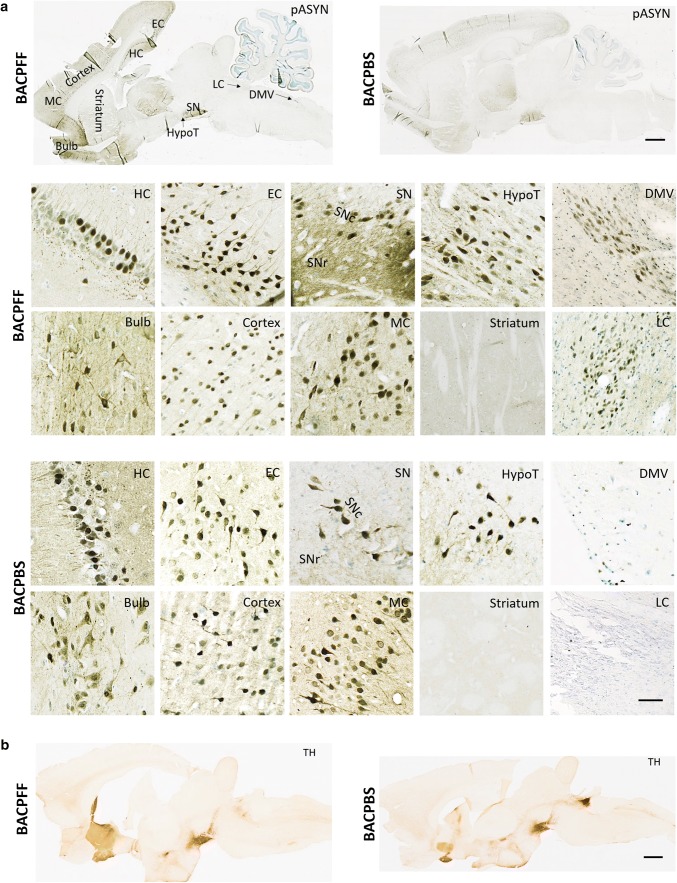
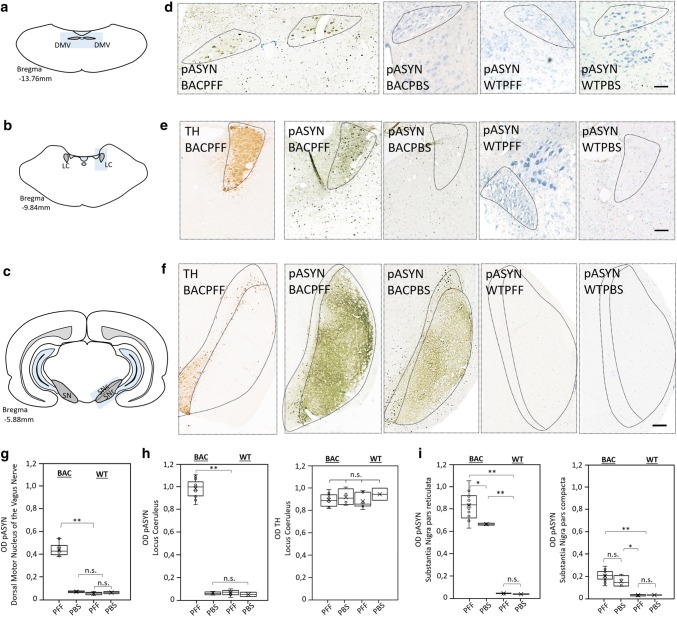
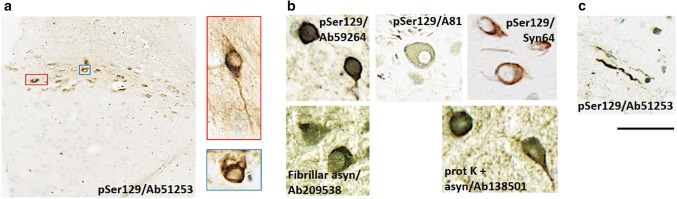
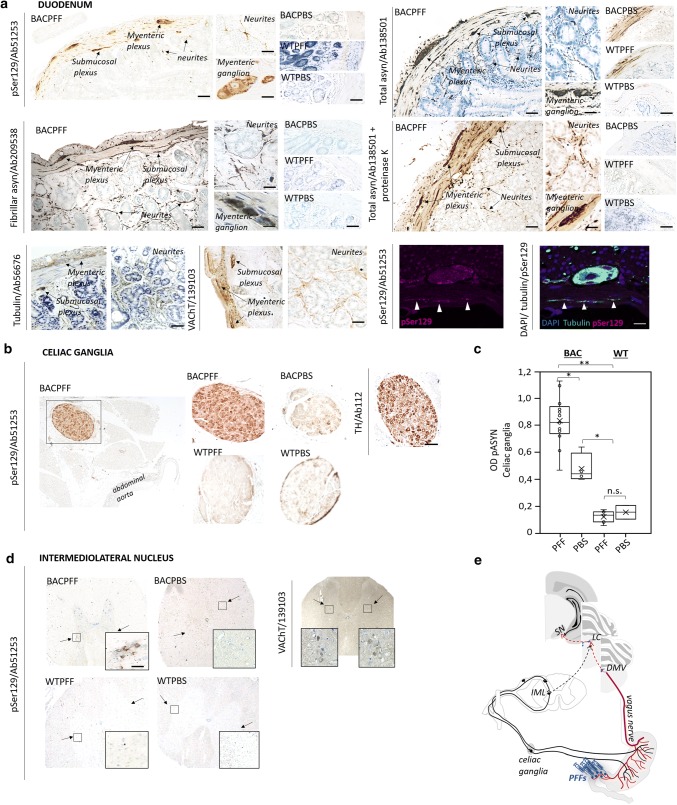
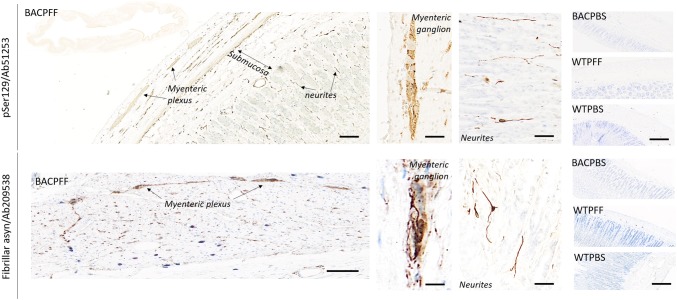
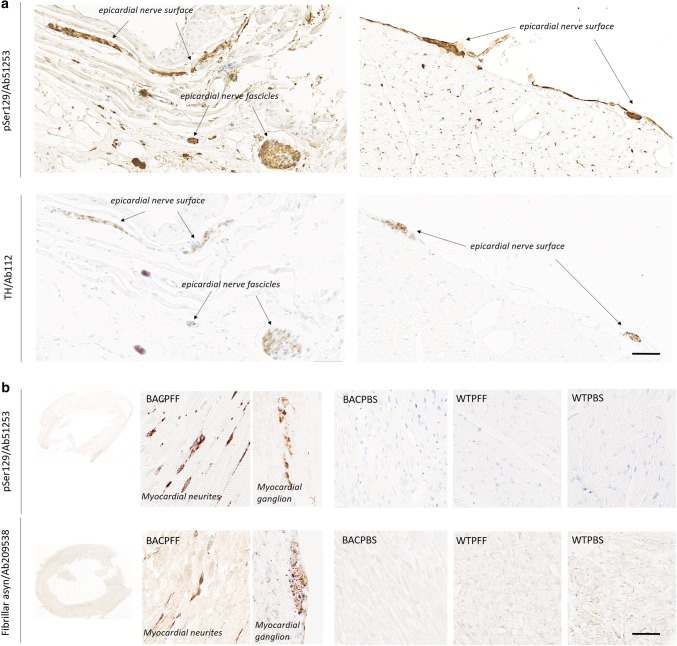
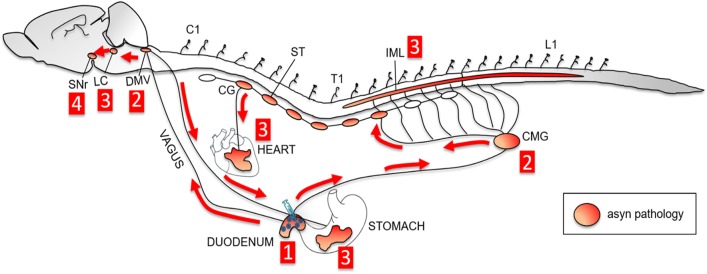
The conversion of endogenous alpha-synuclein (asyn) to pathological asyn-enriched aggregates is a hallmark of Parkinson's disease (PD). These inclusions can be detected in the central and enteric nervous system (ENS). Moreover, gastrointestinal symptoms can appear up to 20 years before the diagnosis of PD. The dual-hit hypothesis posits that pathological asyn aggregation starts in the ENS, and retrogradely spreads to the brain. In this study, we tested this hypothesis by directly injecting preformed asyn fibrils into the duodenum wall of wild-type rats and transgenic rats with excess levels of human asyn. We provide a meticulous characterization of the bacterial artificial chromosome (BAC) transgenic rat model with respect to initial propagation of pathological asyn along the parasympathetic and sympathetic pathways to the brainstem, by performing immunohistochemistry at early time points post-injection. Induced pathology was observed in all key structures along the sympathetic and parasympathetic pathways (ENS, autonomic ganglia, intermediolateral nucleus of the spinal cord (IML), heart, dorsal motor nucleus of the vagus, and locus coeruleus (LC)) and persisted for at least 4 months post-injection. In contrast, asyn propagation was not detected in wild-type rats, nor in vehicle-injected BAC rats. The presence of pathology in the IML, LC, and heart indicate trans-synaptic spread of the pathology. Additionally, the observed asyn inclusions in the stomach and heart may indicate secondary anterograde propagation after initial retrograde spreading. In summary, trans-synaptic propagation of asyn in the BAC rat model is fully compatible with the "body-first hypothesis" of PD etiopathogenesis. To our knowledge, this is the first animal model evidence of asyn propagation to the heart, and the first indication of bidirectional asyn propagation via the vagus nerve, i.e., duodenum-to-brainstem-to-stomach. The BAC rat model could be very valuable for detailed mechanistic studies of the dual-hit hypothesis, and for studies of disease modifying therapies targeting early pathology in the gastrointestinal tract.
Keywords: Alpha-synuclein; Autonomic nervous system; BAC rat model; Parkinson’s disease; Prion-like spread.
Conflict of interest statement
The author declares that they have no competing interests.
Figures
Comment in
-
New models show gut-brain transmission of Parkinson disease pathology.Nat Rev Neurol. 2019 Sep;15(9):491. doi: 10.1038/s41582-019-0241-x. Nat Rev Neurol. 2019. PMID: 31312039 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
