

The influence of masseter and temporalis sarcomere length operating ranges as determined by laser diffraction on architectural estimates of muscle force and excursion in macaques (Macaca fascicularis and Macaca mulatta)
- PMID: 31254839
- PMCID: PMC6739116
- DOI: 10.1016/j.archoralbio.2019.05.015
The influence of masseter and temporalis sarcomere length operating ranges as determined by laser diffraction on architectural estimates of muscle force and excursion in macaques (Macaca fascicularis and Macaca mulatta)
Abstract
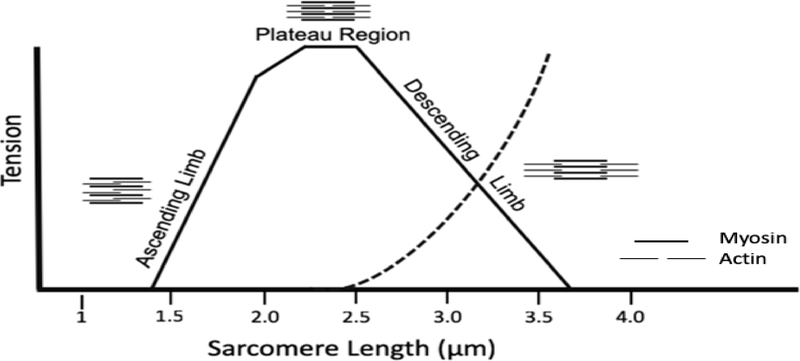
Objective: Determine sarcomere length (Ls) operating ranges of the superficial masseter and temporalis in vitro in a macaque model and examine the impact of position-dependent variation on Ls and architectural estimates of muscle function (i.e., fiber length, PCSA) before and after Ls-normalization.
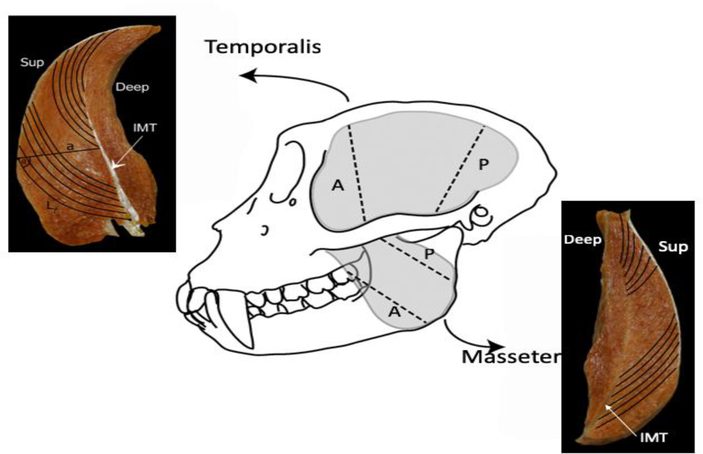
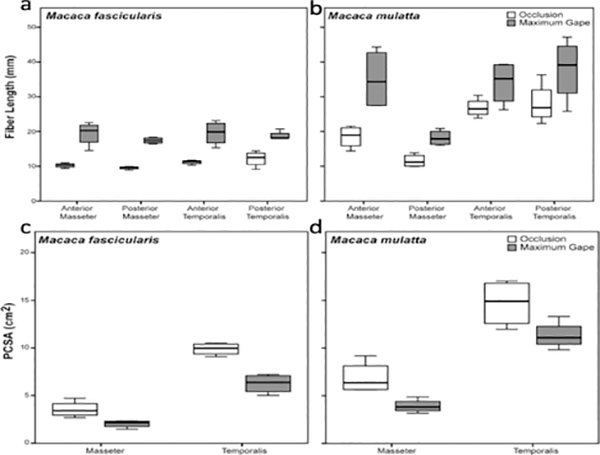
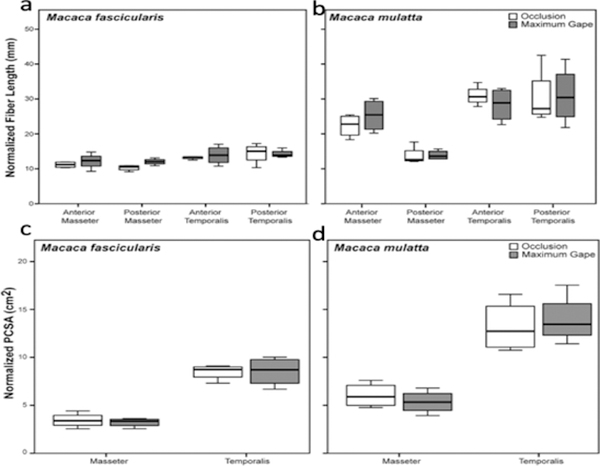
Design: Heads of adult Macaca fascicularis (n = 4) and M. mulatta (n = 3) were bisected postmortem. One side of the jaw was fixed in occlusion, the other in maximum gape. Ls was measured bilaterally using laser diffraction and these measurements were used to estimate sarcomere-length operating ranges. Differences in fiber length and PCSA between sides were tested for significance prior to and following Ls-normalization.
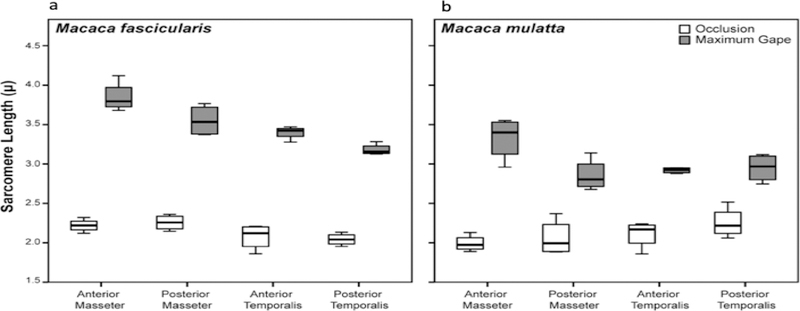
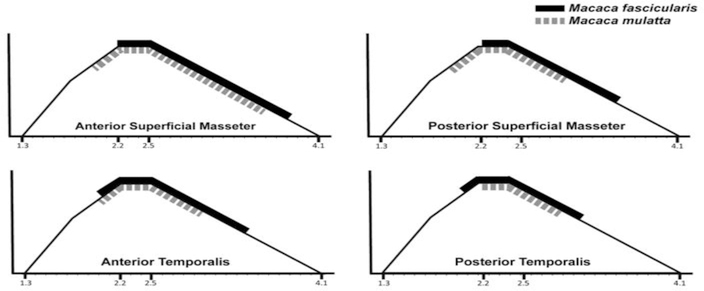
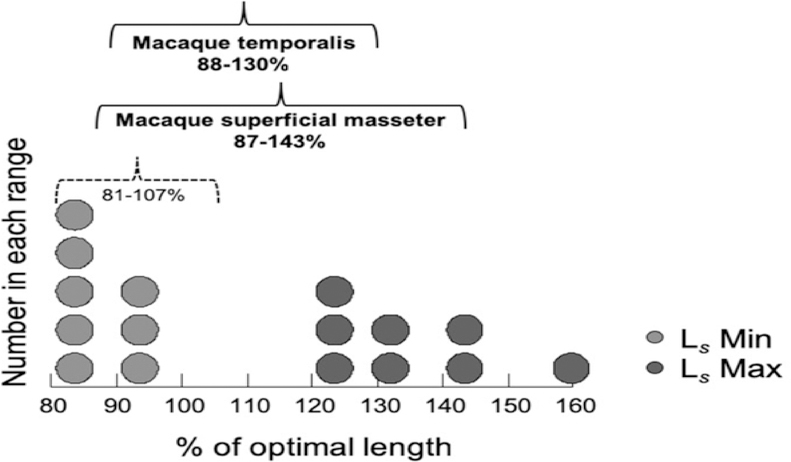
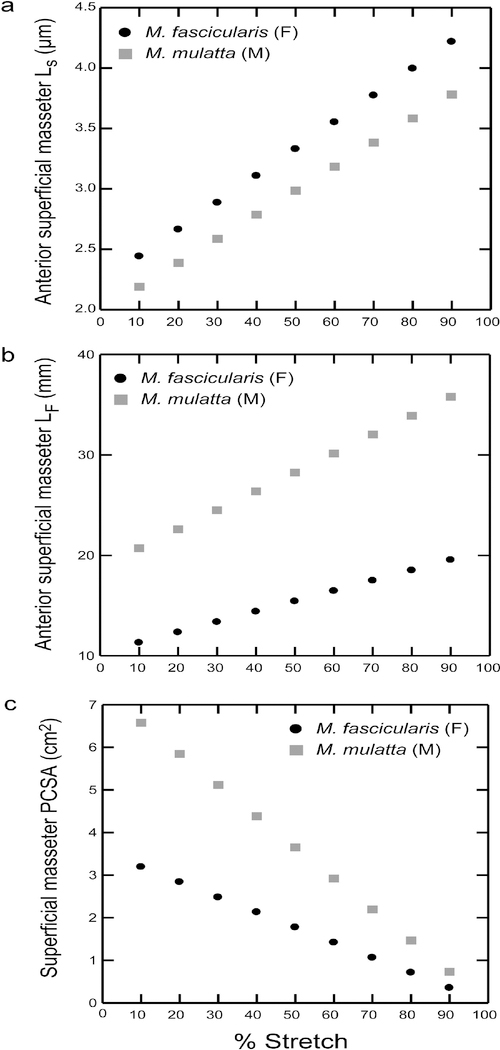
Results: Sarcomere-length operating ranges were widest for the anterior superficial masseter and narrowest for the posterior temporalis. Compared with other mammals, macaque operating ranges were wider and shifted to the right of the descending limb of a representative length-tension curve. Fibers were significantly stretched by as much as 100%, and PCSAs reduced by as much as 43%, on the maximally gaped compared with occluded sides. Ls-normalization substantially reduced position-dependent variance.
Conclusions: The superficial masseter ranges between 87-143% and the temporalis between 88-130% of optimal Ls from maximum gape to occlusion, indicating maximum relative Ls for these macaque muscles exceeds the upper end range previously reported for the jaw muscles of smaller mammals. The wider macaque operating ranges may be functionally linked to the propensity for facially prognathic primates to engage in agonistic canine display behaviors that require jaw-muscle stretch to facilitate production of wide jaw gapes.
Keywords: Fiber length; Macaque; Masseter; Sarcomere length; Temporalis.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflict of interest
None declared.
Figures
References
-
- Anapol F, & Barry K (1996). Fiber architecture of the extensors of the hindlimb in semiterrestrial and arboreal guenons. American Journal of Physical Anthropology, 99, 429–447. - PubMed
-
- Anapol F, & Gray JP (2003). Fiber architecture of the intrinsic muscles of the shoulder and arm in semiterrestrial and arboreal guenons. American Journal of Physical Anthropology, 122, 51–65. - PubMed
-
- Anapol F, & Jungers WL (1986). Architectural and histochemical diversity within the quadriceps femoris of the brown lemur (Lemur fulvus). American Journal of Physical Anthropology, 69, 355–375. - PubMed
-
- Anapol F, Shahnoor N, & Gray JP (2004). Fiber architecture, muscle function, and behavior: Gluteal and hamstring muscles of semiterrestrial and arboreal guenons In Anapol F, German RZ & Jablonski NG (Eds.), Shaping Primate Evolution (pp. 99–133). Cambridge: Cambridge Studies in Biological and Evolutionary Anthropology.
-
- Anapol F, Shahnoor N, & Ross CF (2008). Scaling of reduced physiologic cross-sectional area in primate muscles of mastication In Vinyard CJ, Ravosa MJ & Wall CE (Eds.), Primate Craniofacial Function and Biology (pp. 201–215). New York: Springer.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
