

Bmi1 Suppresses Adipogenesis in the Hematopoietic Stem Cell Niche
- PMID: 31257132
- PMCID: PMC6739622
- DOI: 10.1016/j.stemcr.2019.05.027
Bmi1 Suppresses Adipogenesis in the Hematopoietic Stem Cell Niche
Abstract
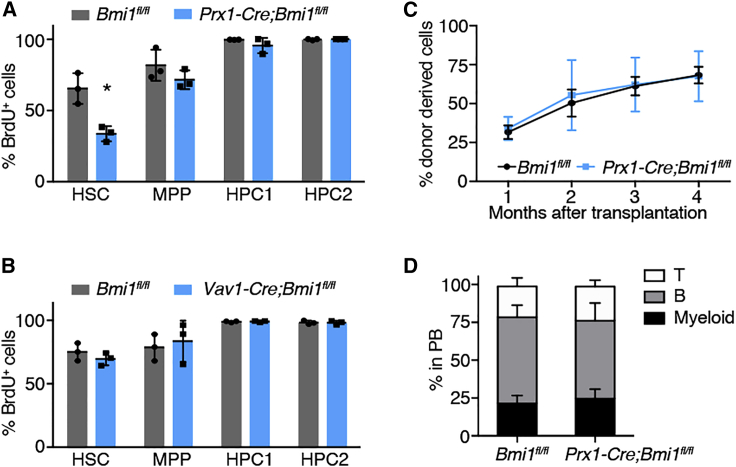
Bone marrow stromal cells (BMSCs) that express high levels of stem cell factor (SCF) and CXC chemokine ligand 12 (CXCL12) are one crucial component of the hematopoietic stem cell (HSC) niche. While the secreted factors produced by BMSCs to support HSCs have been well described, little is known regarding the transcriptional regulators controlling the cell fate of BMSCs and thus indirectly maintaining HSCs. BMI1 is a polycomb group protein that regulates HSCs both cell intrinsically and extrinsically, but it is unknown in which cell type and how BMI1 functions to maintain HSCs extrinsically. Here we show that Bmi1 maintains HSCs by preventing adipogenic differentiation of BMSCs. Bmi1 is highly expressed in BMSCs but becomes downregulated upon adipogenic differentiation and during aging. Deleting Bmi1 from BMSCs increased marrow adipocytes, induced HSC quiescence and depletion, and impaired hematopoiesis. We found that BMI1 repressed multiple developmental programs in BMSCs by safeguarding the repressive epigenetic marks histone H2A ubiquitylation and H3 lysine 27 trimethylation. We identified a novel adipogenic program governed by Pax3, which BMI1 repressed in BMSCs. Our results establish Bmi1 as a critical regulator of BMSC cell fate that suppresses marrow adipogenesis to create a supportive niche for HSCs.
Keywords: BMI1; PAX3; adipocytes; epigenetics; hematopoietic stem cells; mesenchymal stromal cells; niche; polycomb.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Figures







References
-
- Alkema M.J., van der Lugt N.M., Bobeldijk R.C., Berns A., van Lohuizen M. Transformation of axial skeleton due to overexpression of bmi-1 in transgenic mice. Nature. 1995;374:724–727. - PubMed
-
- Azuara V., Perry P., Sauer S., Spivakov M., Jorgensen H.F., John R.M., Gouti M., Casanova M., Warnes G., Merkenschlager M. Chromatin signatures of pluripotent cell lines. Nat. Cell Biol. 2006;8:532–538. - PubMed
-
- Bernstein B.E., Mikkelsen T.S., Xie X., Kamal M., Huebert D.J., Cuff J., Fry B., Meissner A., Wernig M., Plath K. A bivalent chromatin structure marks key developmental genes in embryonic stem cells. Cell. 2006;125:315–326. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
