

Revisiting Trade-offs between Rubisco Kinetic Parameters
- PMID: 31259528
- PMCID: PMC6686151
- DOI: 10.1021/acs.biochem.9b00237
Revisiting Trade-offs between Rubisco Kinetic Parameters
Abstract
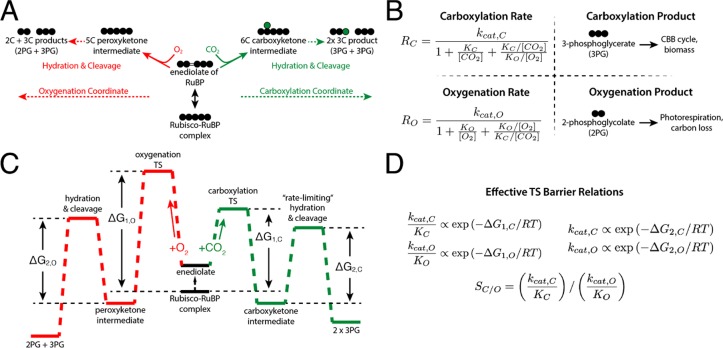
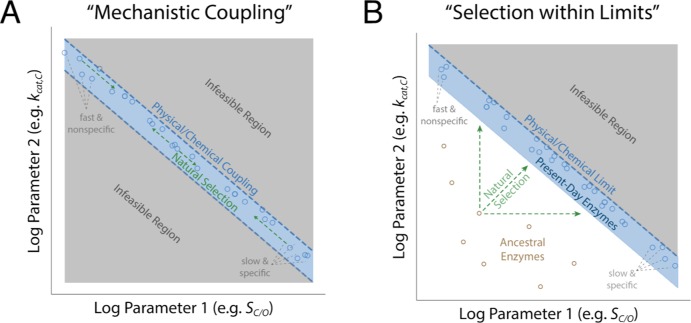
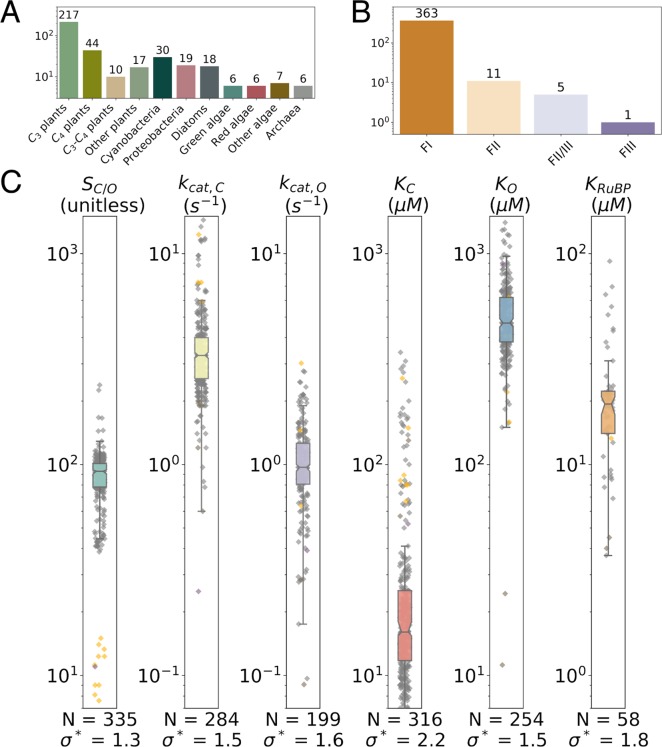
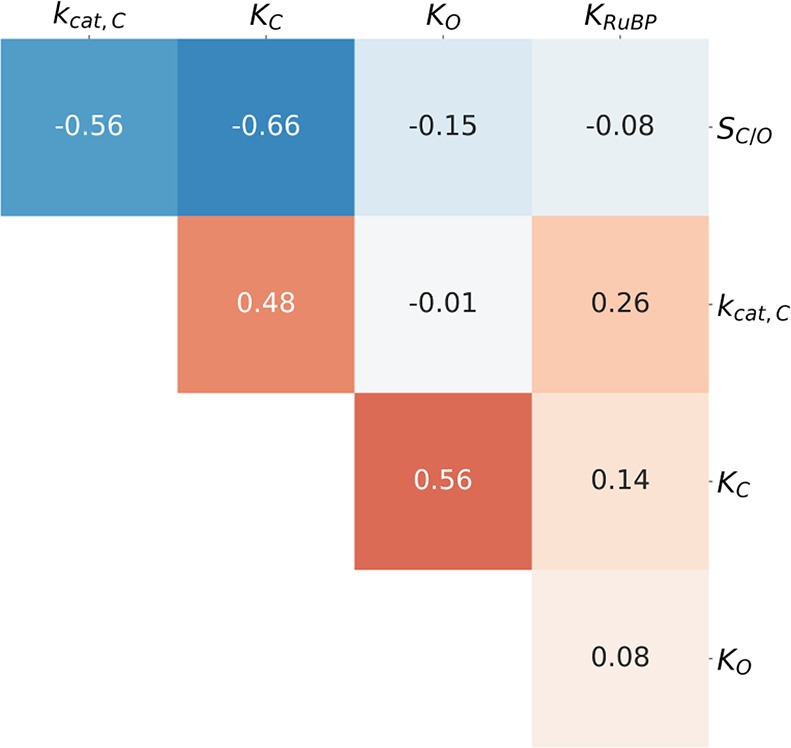
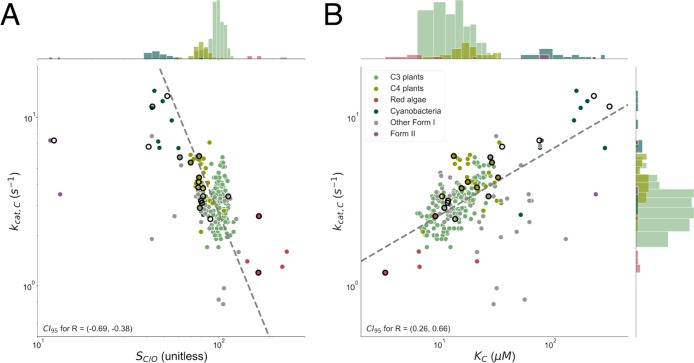
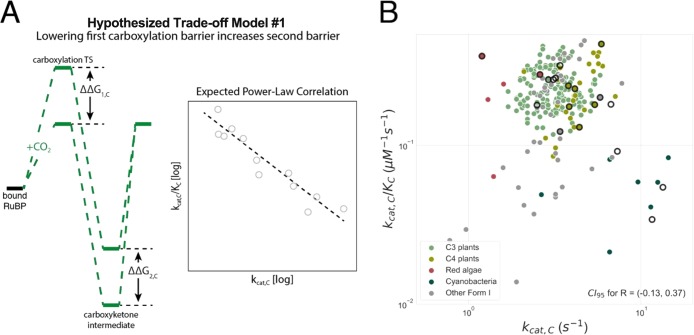
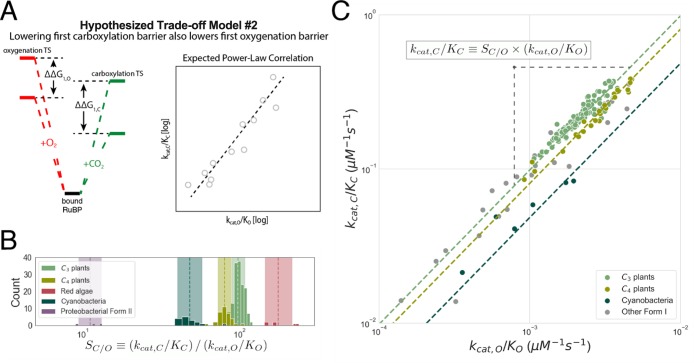
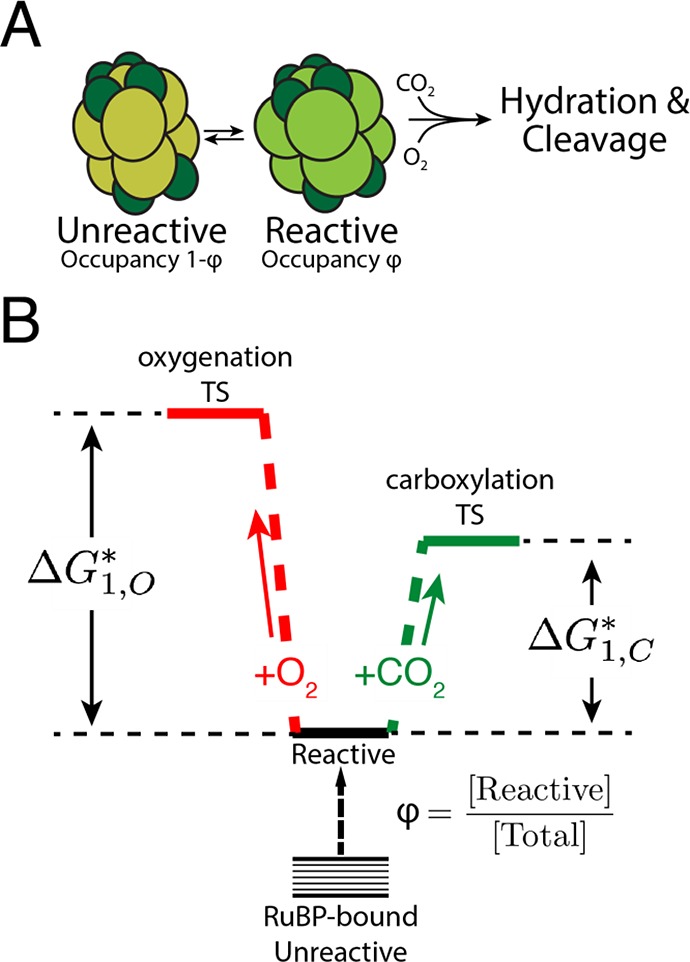
Rubisco is the primary carboxylase of the Calvin cycle, the most abundant enzyme in the biosphere, and one of the best-characterized enzymes. On the basis of correlations between Rubisco kinetic parameters, it is widely posited that constraints embedded in the catalytic mechanism enforce trade-offs between CO2 specificity, SC/O, and maximum carboxylation rate, kcat,C. However, the reasoning that established this view was based on data from ≈20 organisms. Here, we re-examine models of trade-offs in Rubisco catalysis using a data set from ≈300 organisms. Correlations between kinetic parameters are substantially attenuated in this larger data set, with the inverse relationship between kcat,C and SC/O being a key example. Nonetheless, measured kinetic parameters display extremely limited variation, consistent with a view of Rubisco as a highly constrained enzyme. More than 95% of kcat,C values are between 1 and 10 s-1, and no measured kcat,C exceeds 15 s-1. Similarly, SC/O varies by only 30% among Form I Rubiscos and <10% among C3 plant enzymes. Limited variation in SC/O forces a strong positive correlation between the catalytic efficiencies (kcat/KM) for carboxylation and oxygenation, consistent with a model of Rubisco catalysis in which increasing the rate of addition of CO2 to the enzyme-substrate complex requires an equal increase in the O2 addition rate. Altogether, these data suggest that Rubisco evolution is tightly constrained by the physicochemical limits of CO2/O2 discrimination.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
