

Platelet hyaluronidase-2 regulates the early stages of inflammatory disease in colitis
- PMID: 31262781
- PMCID: PMC6716076
- DOI: 10.1182/blood.2018893594
Platelet hyaluronidase-2 regulates the early stages of inflammatory disease in colitis
Abstract
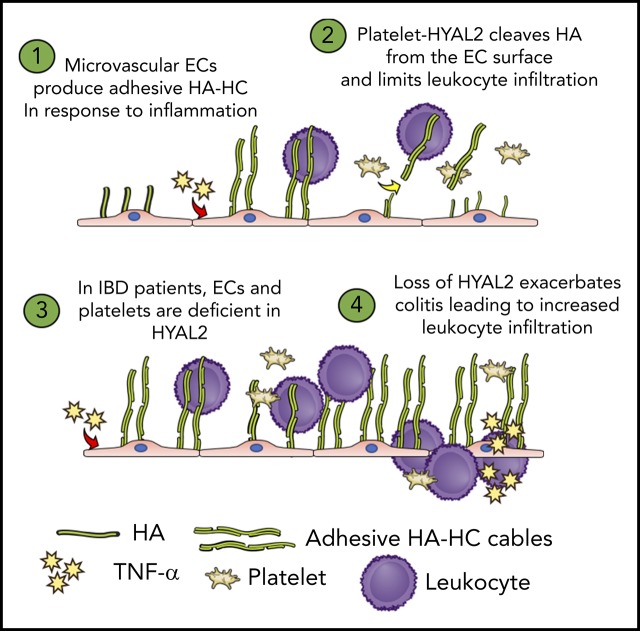
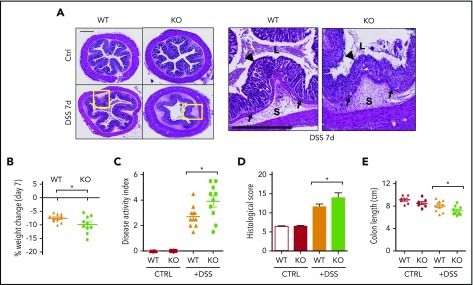
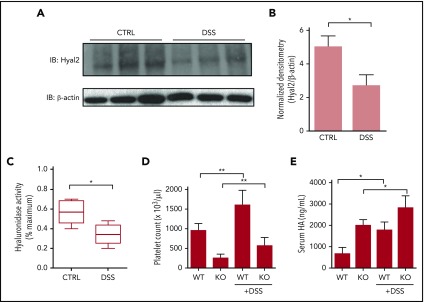
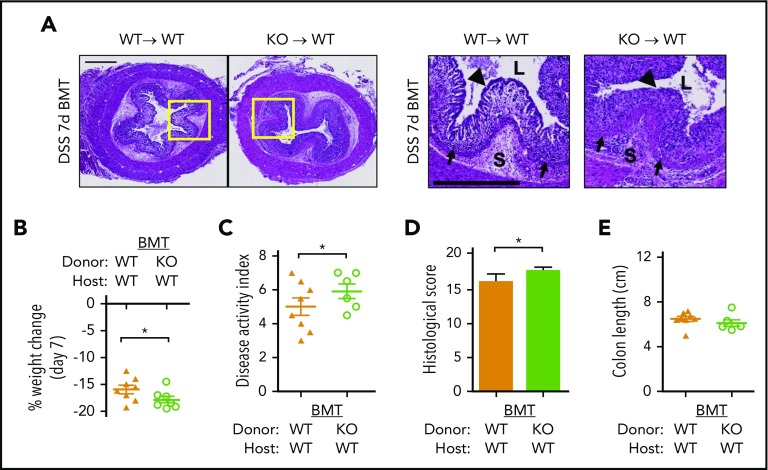
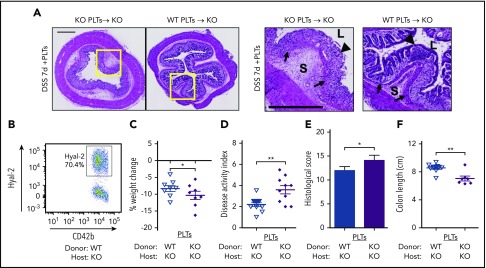
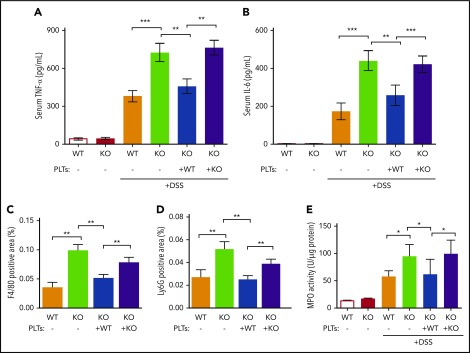
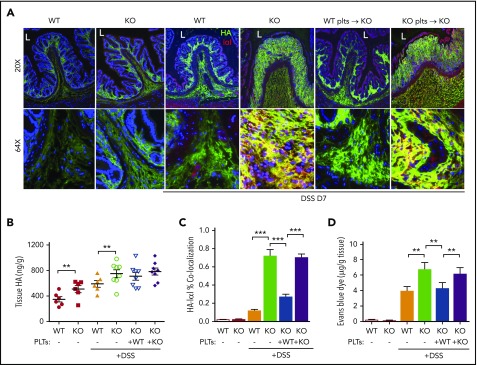
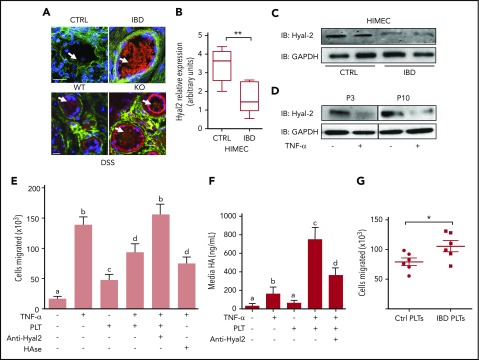
Platelets are specialized cells essential for hemostasis that also function as crucial effectors capable of mediating inflammatory and immune responses. These sentinels continually survey their environment and discriminate between homeostatic and danger signals such as modified components of the extracellular matrix. The glycosaminoglycan hyaluronan (HA) is a major extracellular matrix component that coats the vascular lumen and, under normal conditions, restricts access of inflammatory cells. In response to tissue damage, the endothelial HA matrix enhances leukocyte recruitment and regulates the early stages of the inflammatory response. We have shown that platelets can degrade HA from the surface of activated endothelial cells via the enzyme hyaluronidase-2 (HYAL2) and that HYAL2 is deficient in platelets isolated from patients with inflammatory bowel disease (IBD). Platelets are known to be involved in the pathogenesis of several chronic disease states, including IBD, but they have been largely overlooked in the context of intestinal inflammation. We therefore wanted to define the mechanism by which platelet HYAL2 regulates the inflammatory response during colitis. In this study, we provide evidence that HA catabolism is disrupted in human intestinal microvascular endothelial cells isolated from patients with IBD. Furthermore, mice deficient in HYAL2 are more susceptible to an acute model of colitis, and this increased susceptibility is abrogated by transfusion of HYAL2-competent platelets. Finally, we show that platelets, via HYAL2-dependent degradation of endothelial HA, regulate the early stages of inflammation in colitis by limiting leukocyte extravasation.
© 2019 by The American Society of Hematology.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
References
-
- de Souza HS, Fiocchi C. Immunopathogenesis of IBD: current state of the art. Nat Rev Gastroenterol Hepatol. 2016;13(1):13-27. - PubMed
-
- Lees CW, Barrett JC, Parkes M, Satsangi J. New IBD genetics: common pathways with other diseases. Gut. 2011;60(12):1739-1753. - PubMed
-
- Fiocchi C. Intestinal inflammation: a complex interplay of immune and nonimmune cell interactions. Am J Physiol. 1997;273(4):G769-G775. - PubMed
-
- Danese S, Motte Cd CL, Fiocchi C. Platelets in inflammatory bowel disease: clinical, pathogenic, and therapeutic implications. Am J Gastroenterol. 2004;99(5):938-945. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
