

Mapping the native organization of the yeast nuclear pore complex using nuclear radial intensity measurements
- PMID: 31262825
- PMCID: PMC6642398
- DOI: 10.1073/pnas.1903764116
Mapping the native organization of the yeast nuclear pore complex using nuclear radial intensity measurements
Abstract
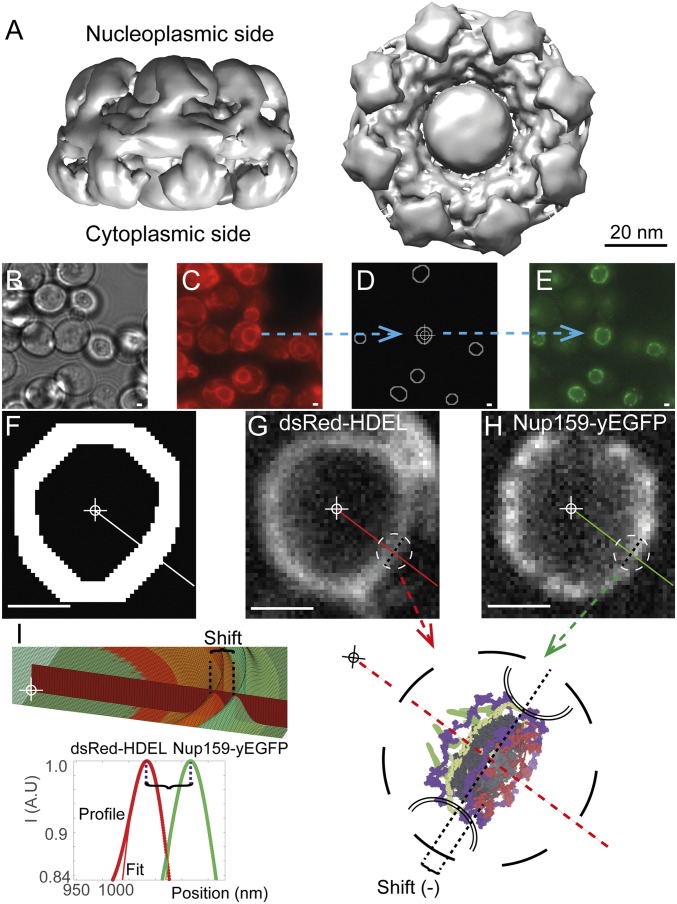
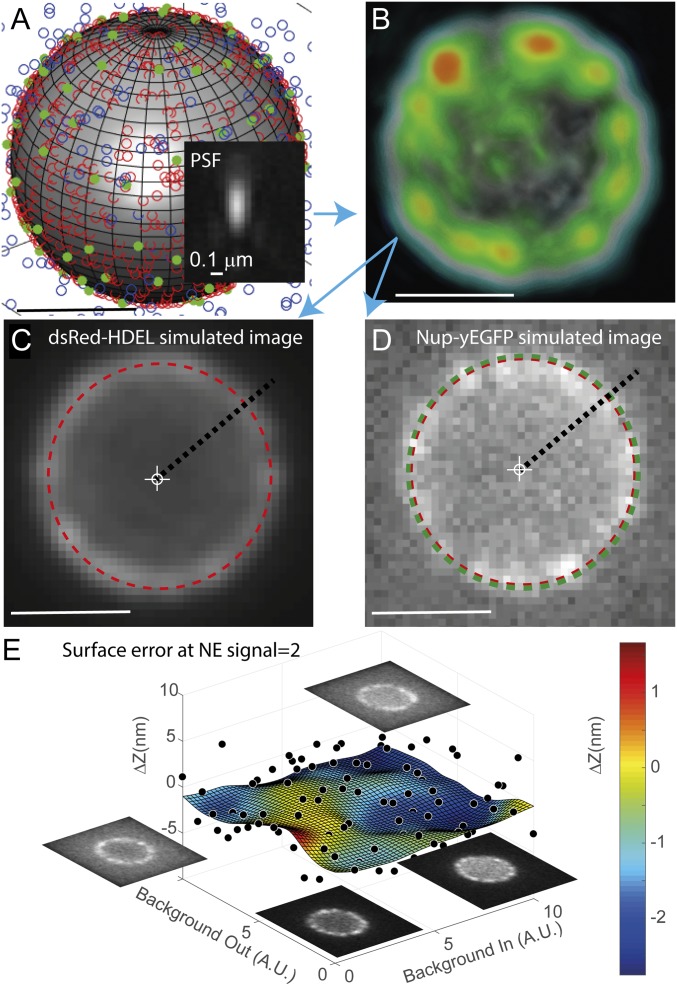
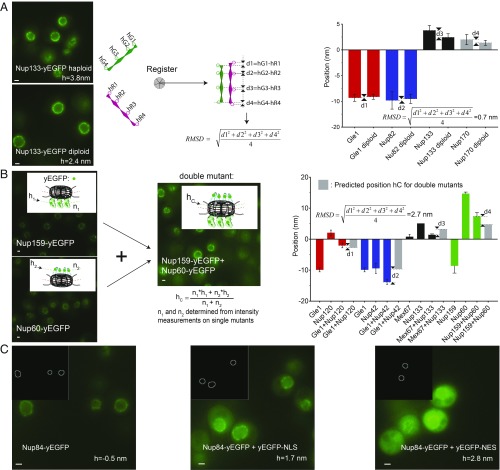
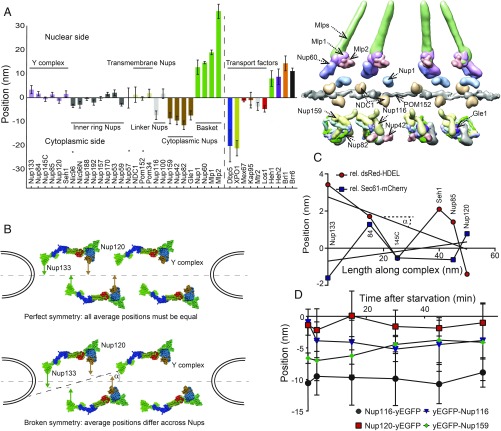
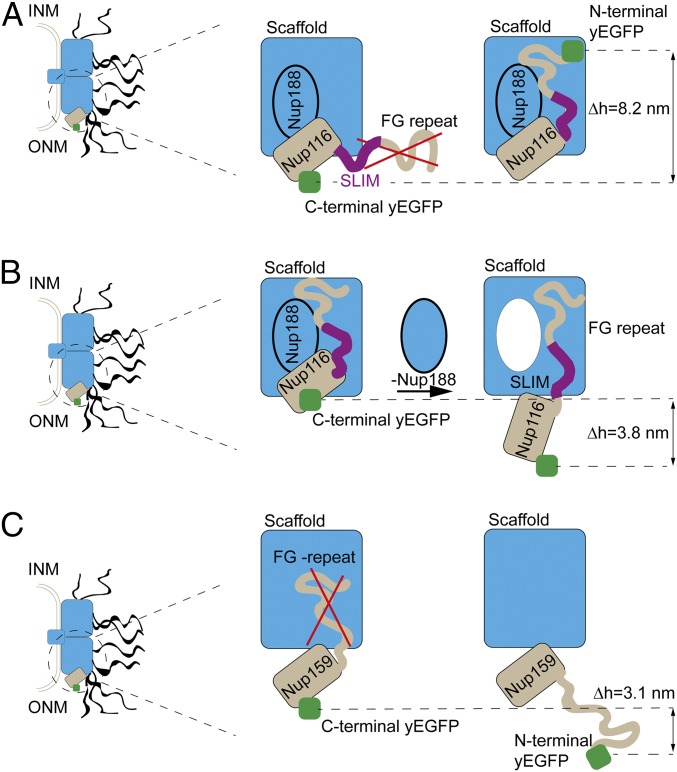
Selective transport across the nuclear envelope (NE) is mediated by the nuclear pore complex (NPC), a massive ∼100-MDa assembly composed of multiple copies of ∼30 nuclear pore proteins (Nups). Recent advances have shed light on the composition and structure of NPCs, but approaches that could map their organization in live cells are still lacking. Here, we introduce an in vivo method to perform nuclear radial intensity measurements (NuRIM) using fluorescence microscopy to determine the average position of NE-localized proteins along the nucleocytoplasmic transport axis. We apply NuRIM to study the organization of the NPC and the mobile transport machinery in budding yeast. This reveals a unique snapshot of the intact yeast NPC and identifies distinct steady-state localizations for various NE-associated proteins and nuclear transport factors. We find that the NPC architecture is robust against compositional changes and could also confirm that in contrast to Chlamydomonas reinhardtii, the scaffold Y complex is arranged symmetrically in the yeast NPC. Furthermore, NuRIM was applied to probe the orientation of intrinsically disordered FG-repeat segments, providing insight into their roles in selective NPC permeability and structure.
Keywords: FG repeats; nuclear pore complex; nucleoporins; quantitative fluorescence microscopy; superresolution.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
