

Coexisting representations of sensory and mnemonic information in human visual cortex
- PMID: 31263205
- PMCID: PMC6857532
- DOI: 10.1038/s41593-019-0428-x
Coexisting representations of sensory and mnemonic information in human visual cortex
Abstract
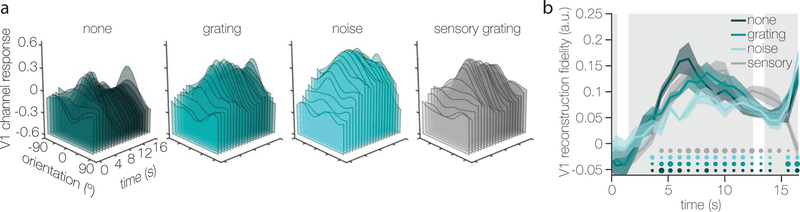
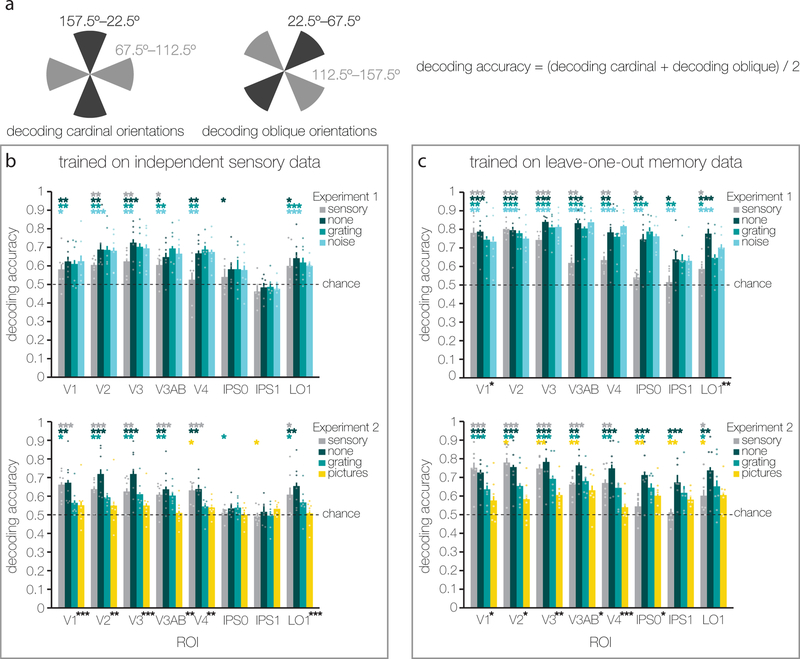
Traversing sensory environments requires keeping relevant information in mind while simultaneously processing new inputs. Visual information is kept in working memory via feature-selective responses in early visual cortex, but recent work has suggested that new sensory inputs obligatorily wipe out this information. Here we show region-wide multiplexing abilities in classic sensory areas, with population-level response patterns in early visual cortex representing the contents of working memory alongside new sensory inputs. In a second experiment, we show that when people get distracted, this leads to both disruptions of mnemonic information in early visual cortex and decrements in behavioral recall. Representations in the intraparietal sulcus reflect actively remembered information encoded in a transformed format, but not task-irrelevant sensory inputs. Together, these results suggest that early visual areas play a key role in supporting high-resolution working memory representations that can serve as a template for comparison with incoming sensory information.
Conflict of interest statement
Figures



References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
