

PGC-1a integrates a metabolism and growth network linked to caloric restriction
- PMID: 31267675
- PMCID: PMC6718593
- DOI: 10.1111/acel.12999
PGC-1a integrates a metabolism and growth network linked to caloric restriction
Abstract
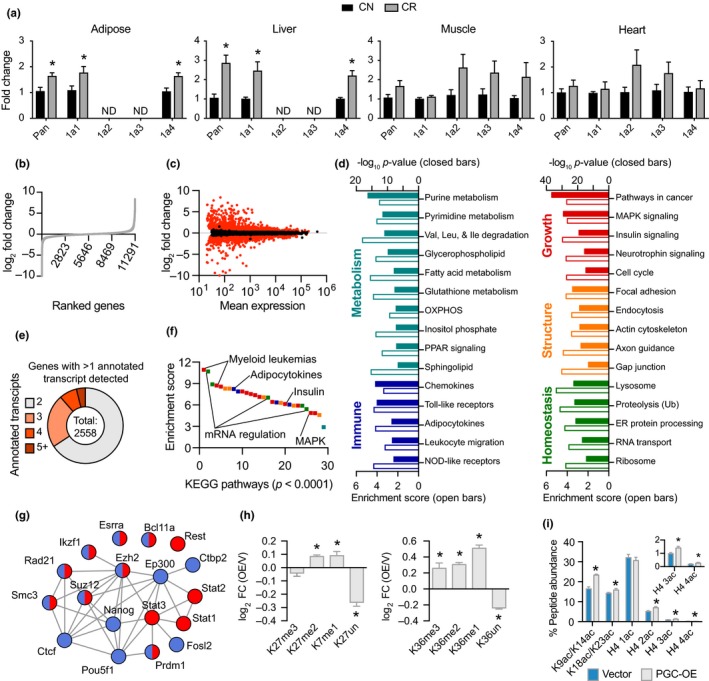
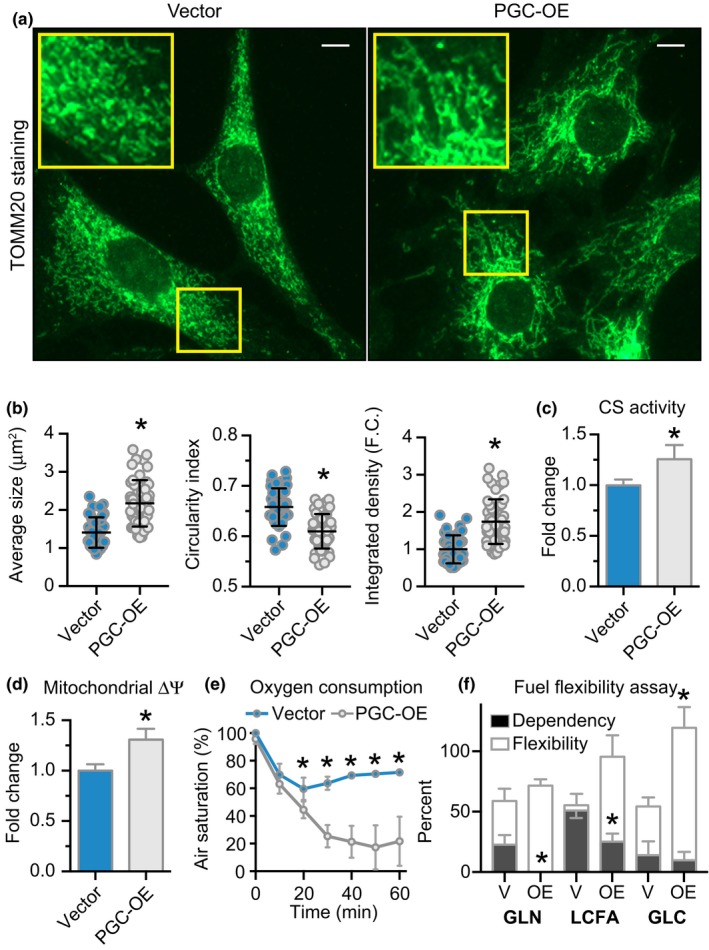
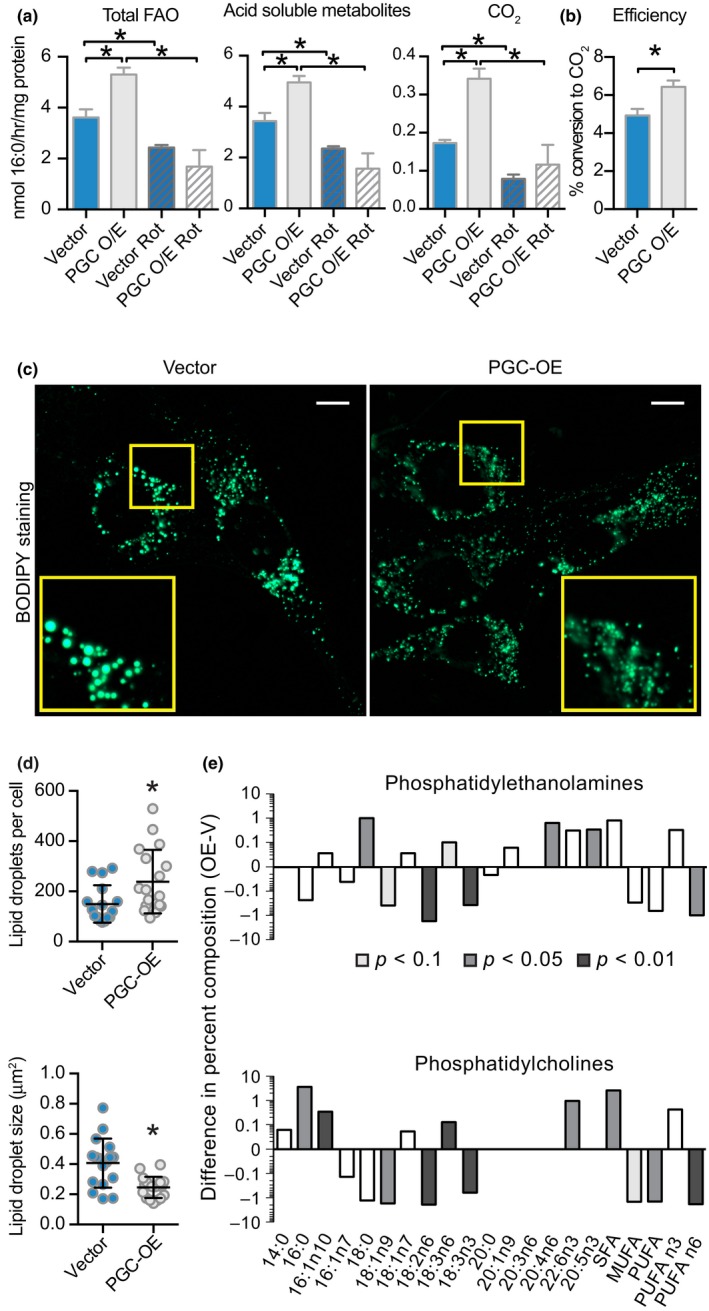
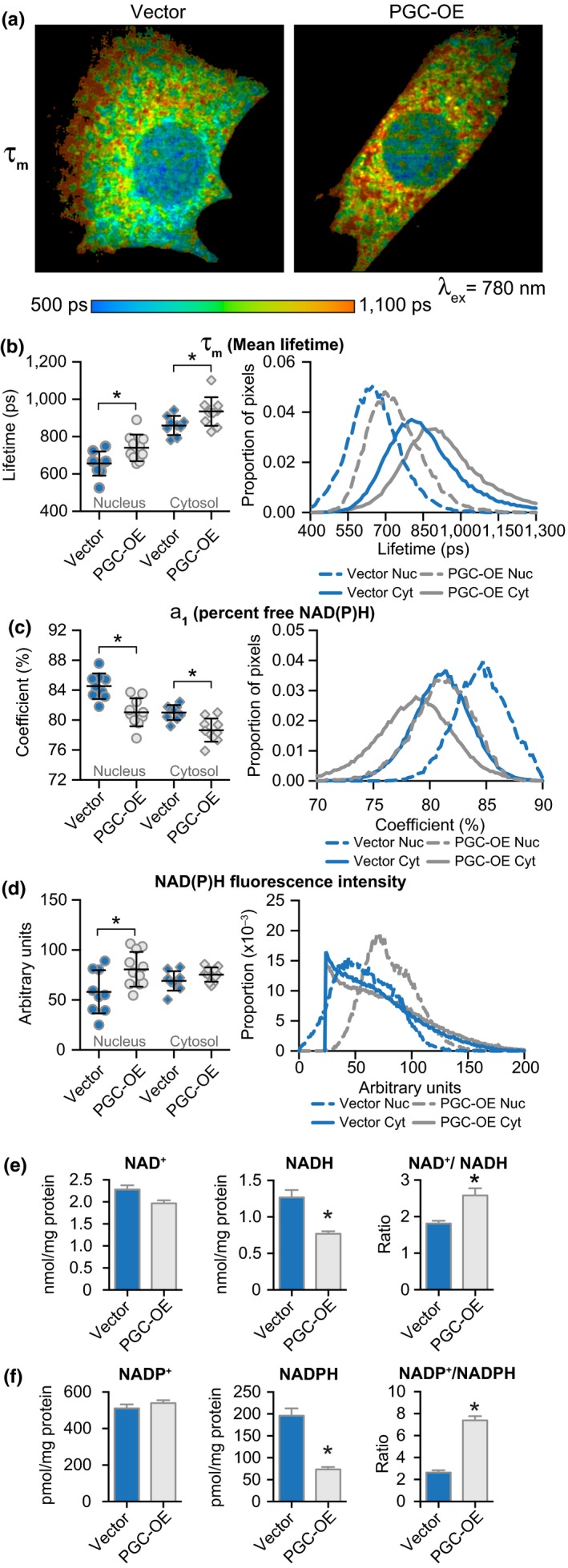
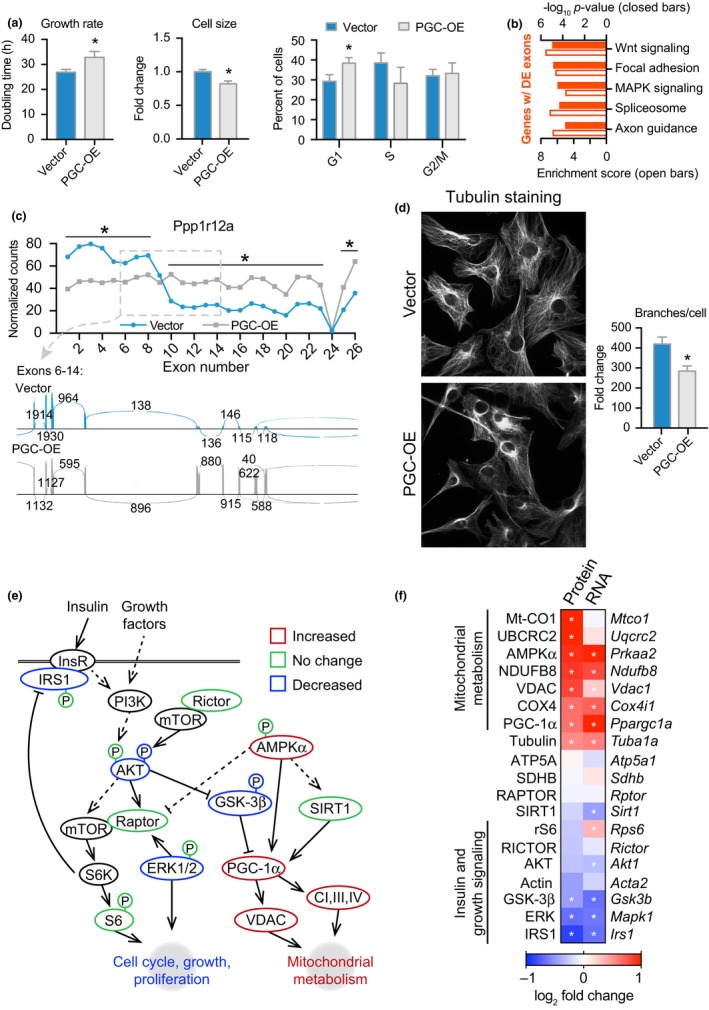
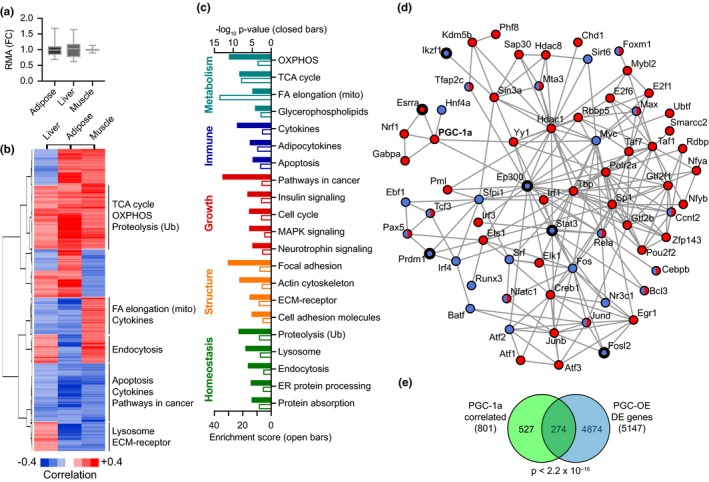
Deleterious changes in energy metabolism have been linked to aging and disease vulnerability, while activation of mitochondrial pathways has been linked to delayed aging by caloric restriction (CR). The basis for these associations is poorly understood, and the scope of impact of mitochondrial activation on cellular function has yet to be defined. Here, we show that mitochondrial regulator PGC-1a is induced by CR in multiple tissues, and at the cellular level, CR-like activation of PGC-1a impacts a network that integrates mitochondrial status with metabolism and growth parameters. Transcriptional profiling reveals that diverse functions, including immune pathways, growth, structure, and macromolecule homeostasis, are responsive to PGC-1a. Mechanistically, these changes in gene expression were linked to chromatin remodeling and RNA processing. Metabolic changes implicated in the transcriptional data were confirmed functionally including shifts in NAD metabolism, lipid metabolism, and membrane lipid composition. Delayed cellular proliferation, altered cytoskeleton, and attenuated growth signaling through post-transcriptional and post-translational mechanisms were also identified as outcomes of PGC-1a-directed mitochondrial activation. Furthermore, in vivo in tissues from a genetically heterogeneous mouse population, endogenous PGC-1a expression was correlated with this same metabolism and growth network. These data show that small changes in metabolism have broad consequences that arguably would profoundly alter cell function. We suggest that this PGC-1a sensitive network may be the basis for the association between mitochondrial function and aging where small deficiencies precipitate loss of function across a spectrum of cellular activities.
Keywords: NAD; PGC-1a; caloric restriction; lipid metabolism; longevity; mitochondria; redox metabolism.
© 2019 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
JMD is a consultant for BioTechne and FORGE Bioscience and is cofounder of Galilei BioSciences. The remaining authors declare no conflict of interest.
Figures
References
-
- Anderson, R. M. , Barger, J. L. , Edwards, M. G. , Braun, K. H. , O'Connor, C. E. , Prolla, T. A. , & Weindruch, R. (2008). Dynamic regulation of PGC‐1alpha localization and turnover implicates mitochondrial adaptation in calorie restriction and the stress response. Aging Cell, 7(1), 101–111. 10.1111/j.1474-9726.2007.00357.x - DOI - PMC - PubMed
-
- Barger, J. L. , Anderson, R. M. , Newton, M. A. , da Silva, C. , Vann, J. A. , Pugh, T. D. , … Weindruch, R. (2015). A conserved transcriptional signature of delayed aging and reduced disease vulnerability is partially mediated by SIRT3. PLoS ONE, 10(4), e0120738 10.1371/journal.pone.0120738 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
