

Vagal innervation of the stomach reassessed: brain-gut connectome uses smart terminals
- PMID: 31268562
- PMCID: PMC6810743
- DOI: 10.1111/nyas.14138
Vagal innervation of the stomach reassessed: brain-gut connectome uses smart terminals
Abstract
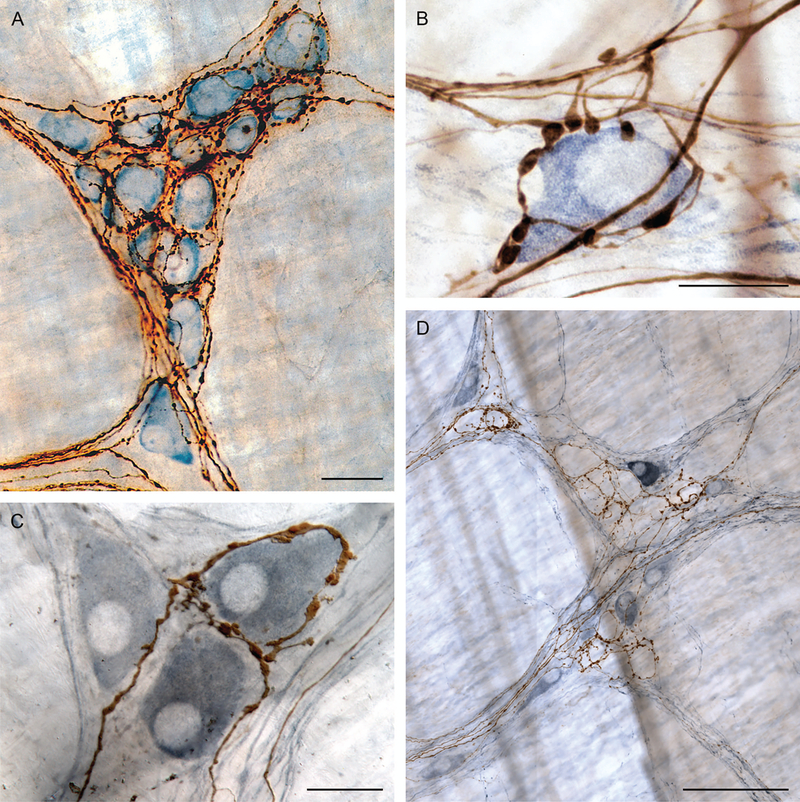
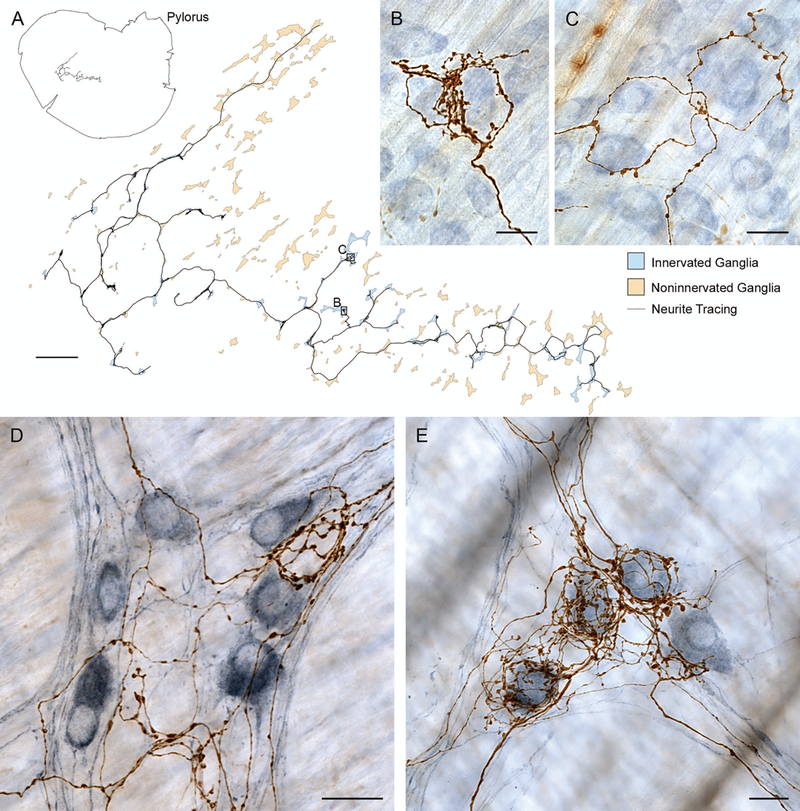
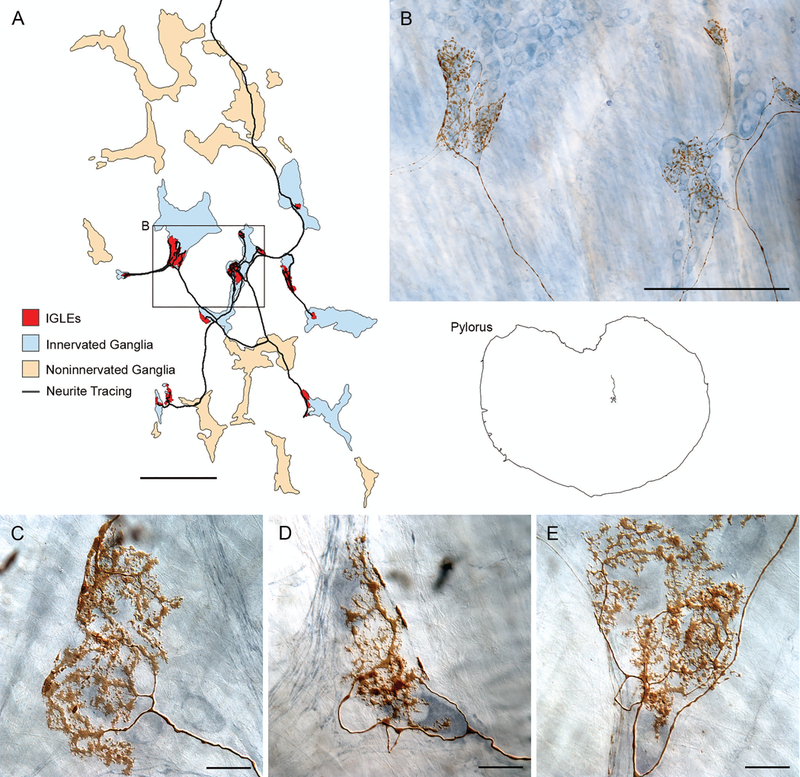
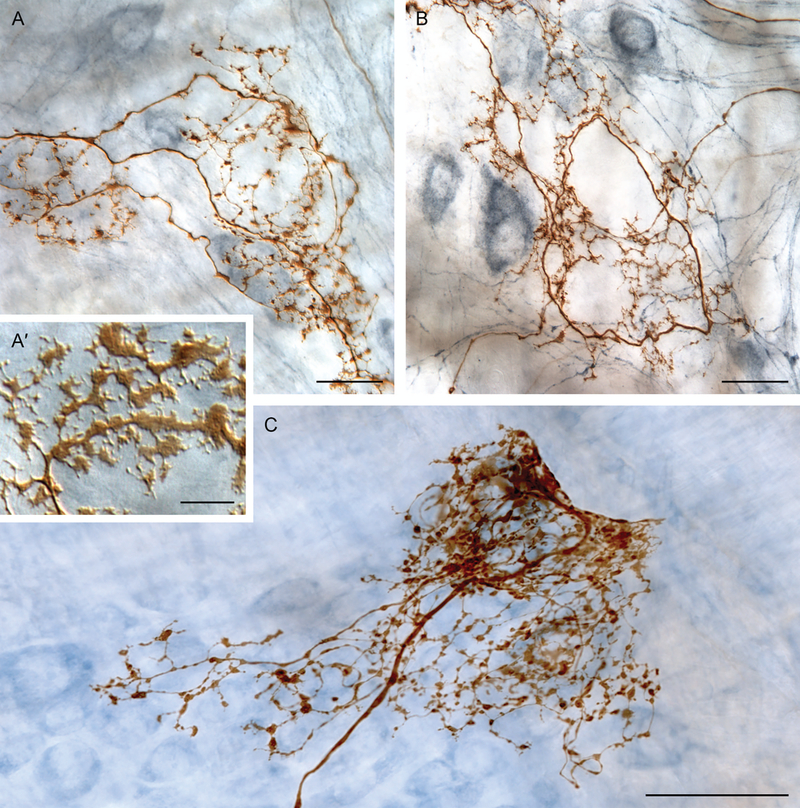
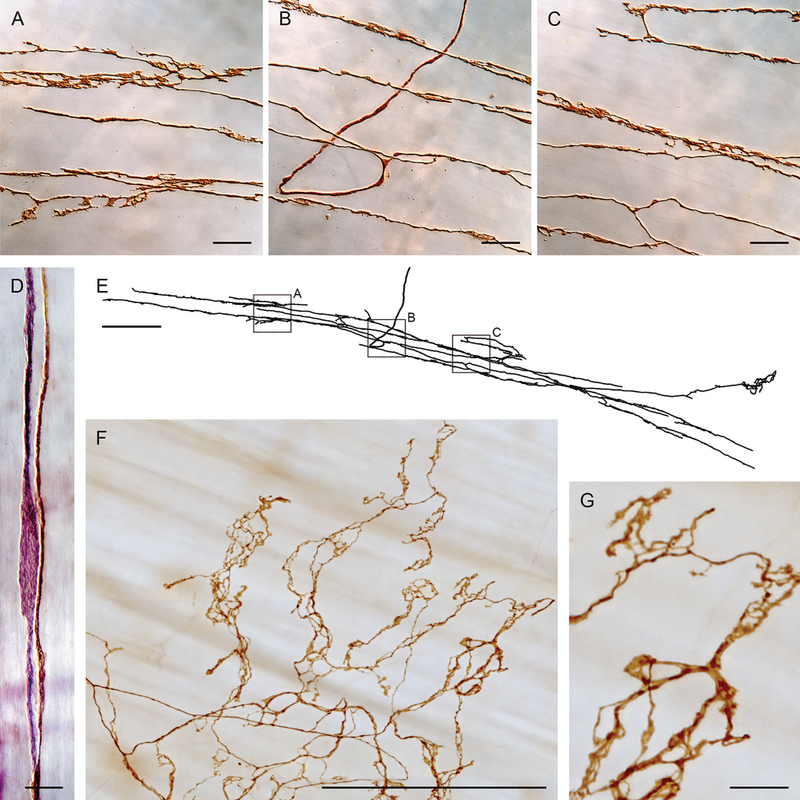
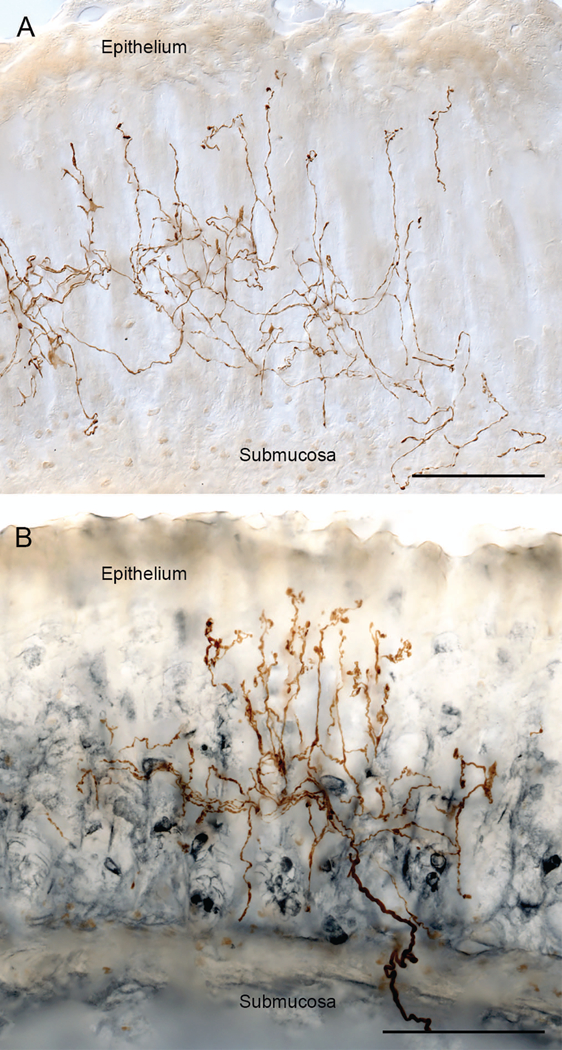
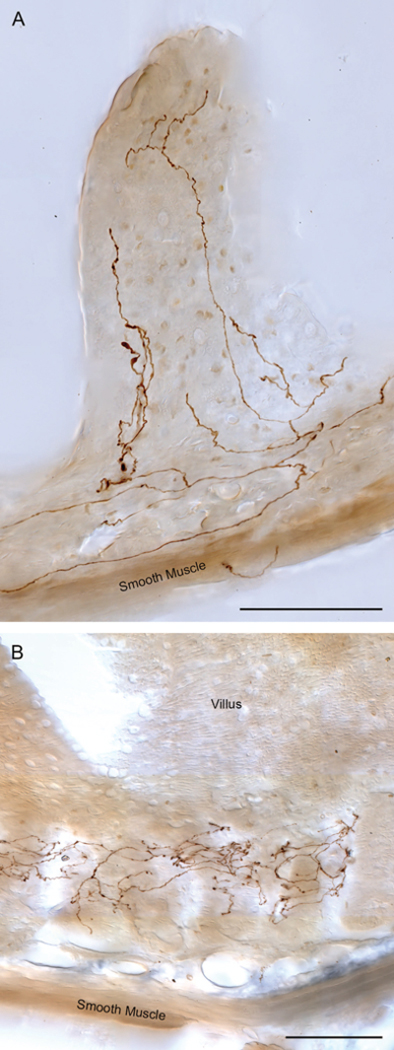
Brain-gut neural communications have long been considered limited because of conspicuous numerical mismatches. The vagus, the parasympathetic nerve connecting brain and gut, contains thousands of axons, whereas the gastrointestinal (GI) tract contains millions of intrinsic neurons in local plexuses. The numerical paradox was initially recognized in terms of efferent projections, but the number of afferents, which comprise the majority (≈ 80%) of neurites in the vagus, is also relatively small. The present survey of recent morphological observations suggests that vagal terminals, and more generally autonomic and visceral afferent arbors in the stomach as well as throughout the gut, elaborate arbors that are extensive, regionally specialized, polymorphic, polytopic, and polymodal, commonly with multiplicities of receptors and binding sites-smart terminals. The morphological specializations and dynamic tuning of one-to-many efferent projections and many-to-one convergences of contacts onto afferents create a complex architecture capable of extensive peripheral integration in the brain-gut connectome and offset many of the disparities between axon and target numbers. Appreciating this complex architecture can help in the design of therapies for GI disorders.
Keywords: afferent; autonomic; efferent; intestines; parasympathetic; postganglionic; preganglionic; stomach; vagus; viscera; visceral afferent.
© 2019 New York Academy of Sciences.
Figures

References
-
- Langley JN 1922. Connexions of the enteric nerve cells. J. Physiol. (Lond.). 56:xxxix
-
- Wood JD 1987. Physiology of the Enteric Nervous System In Physiology of the Gastrointestinal Tract. Johnson LR, Ed.: 2nd ed. New York, Raven Press.
-
- Hill CJ 1927. A contribution to our knowledge of the enteric plexuses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 215:355–387.
-
- Berthoud H-R, Jedrzejewska A, & Powley TL. 1990. Simultaneous labeling of vagal innervation of the gut and afferent projections from the visceral forebrain with Dil injected into the dorsal vagal complex in the rat. J. Comp. Neurol. 301(1):65–79. - PubMed
-
- Holst M-C, Kelly JB & Powley TL. 1997. Vagal preganglionic projections to the enteric nervous system characterized with PHA-L. J. Comp. Neurol. 381:81–100. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
