

Active membrane conductances and morphology of a collision detection neuron broaden its impedance profile and improve discrimination of input synchrony
- PMID: 31268830
- PMCID: PMC6734408
- DOI: 10.1152/jn.00048.2019
Active membrane conductances and morphology of a collision detection neuron broaden its impedance profile and improve discrimination of input synchrony
Abstract
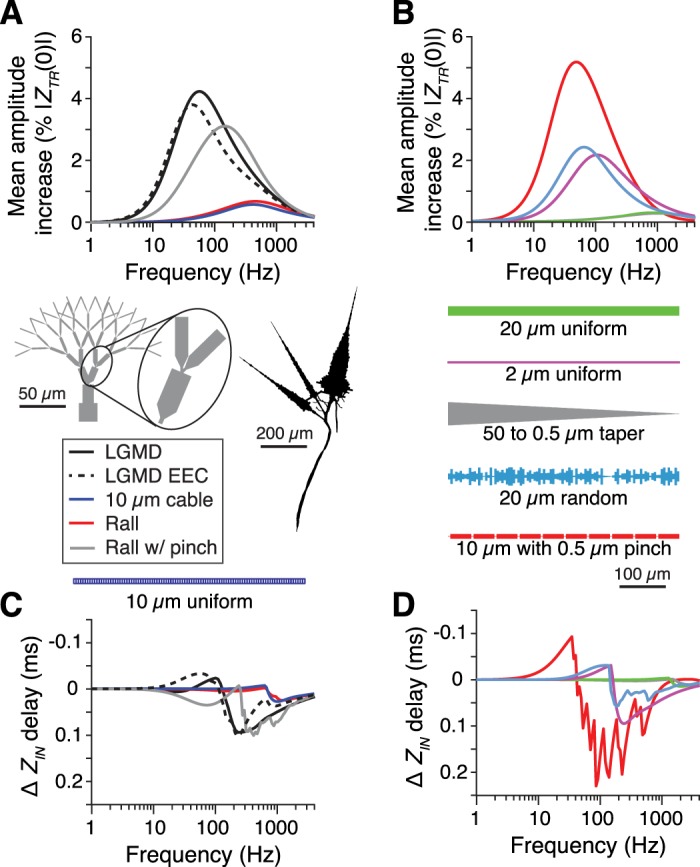
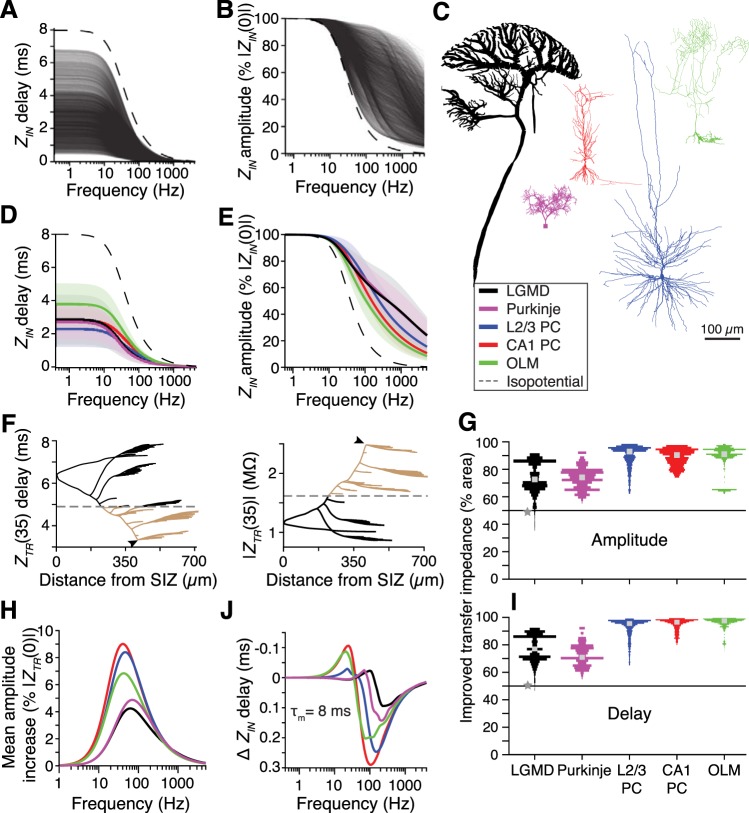
How neurons filter and integrate their complex patterns of synaptic inputs is central to their role in neural information processing. Synaptic filtering and integration are shaped by the frequency-dependent neuronal membrane impedance. Using single and dual dendritic recordings in vivo, pharmacology, and computational modeling, we characterized the membrane impedance of a collision detection neuron in the grasshopper Schistocerca americana. This neuron, the lobula giant movement detector (LGMD), exhibits consistent impedance properties across frequencies and membrane potentials. Two common active conductances gH and gM, mediated respectively by hyperpolarization-activated cyclic nucleotide-gated (HCN) channels and by muscarine-sensitive M-type K+ channels, promote broadband integration with high temporal precision over the LGMD's natural range of membrane potentials and synaptic input frequencies. Additionally, we found that a model based on the LGMD's branching morphology increased the gain and decreased the delay associated with the mapping of synaptic input currents to membrane potential. More generally, this was true for a wide range of model neuron morphologies, including those of neocortical pyramidal neurons and cerebellar Purkinje cells. These findings show the unexpected role played by two widespread active conductances and by dendritic morphology in shaping synaptic integration.NEW & NOTEWORTHY Neuronal filtering and integration of synaptic input patterns depend on the electrochemical properties of dendrites. We used an identified collision detection neuron in grasshoppers to examine how its morphology and two conductances affect its membrane impedance in relation to the computations it performs. The neuronal properties examined are ubiquitous and therefore promote a general understanding of neuronal computations, including those in the human brain.
Keywords: collision avoidance; dendritic processing; lobula giant movement detector; membrane impedance.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases