

GPIHBP1 and Lipoprotein Lipase, Partners in Plasma Triglyceride Metabolism
- PMID: 31269429
- PMCID: PMC6662658
- DOI: 10.1016/j.cmet.2019.05.023
GPIHBP1 and Lipoprotein Lipase, Partners in Plasma Triglyceride Metabolism
Abstract
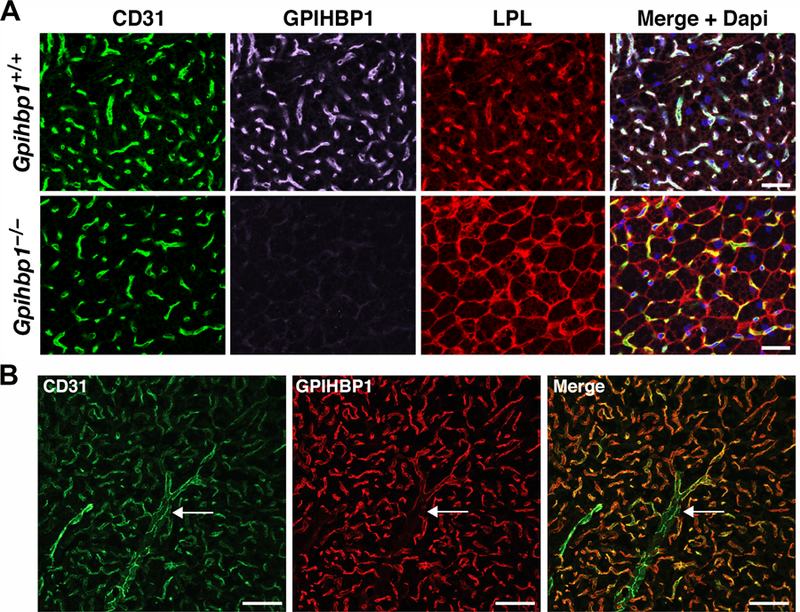
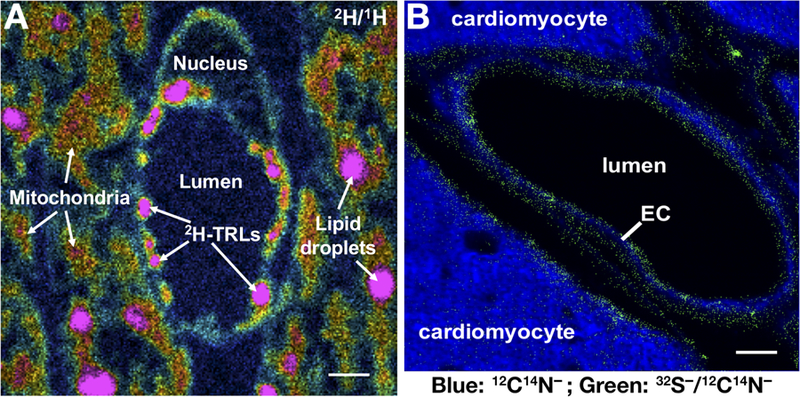
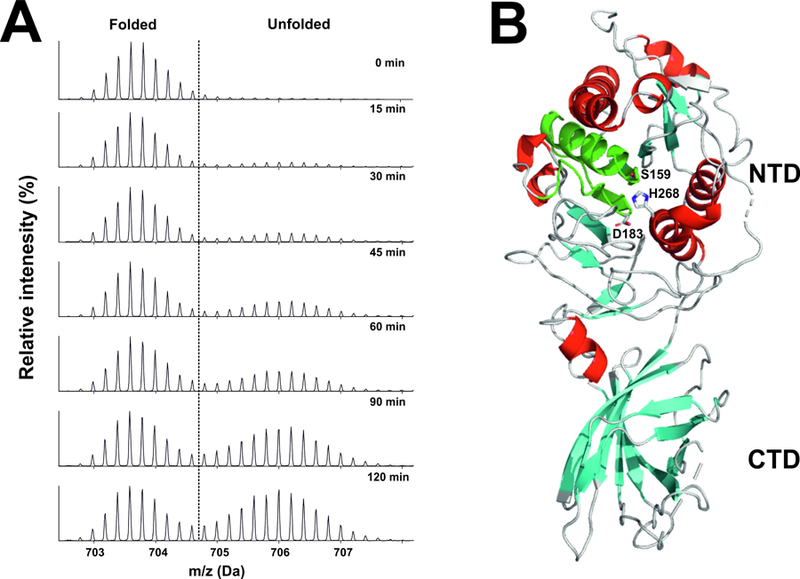
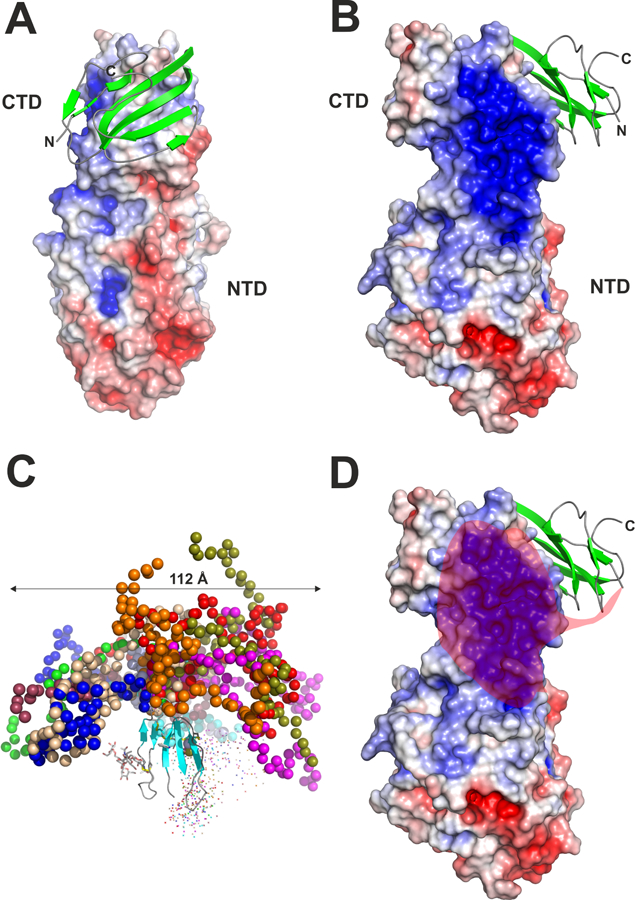
Lipoprotein lipase (LPL), identified in the 1950s, has been studied intensively by biochemists, physiologists, and clinical investigators. These efforts uncovered a central role for LPL in plasma triglyceride metabolism and identified LPL mutations as a cause of hypertriglyceridemia. By the 1990s, with an outline for plasma triglyceride metabolism established, interest in triglyceride metabolism waned. In recent years, however, interest in plasma triglyceride metabolism has awakened, in part because of the discovery of new molecules governing triglyceride metabolism. One such protein-and the focus of this review-is GPIHBP1, a protein of capillary endothelial cells. GPIHBP1 is LPL's essential partner: it binds LPL and transports it to the capillary lumen; it is essential for lipoprotein margination along capillaries, allowing lipolysis to proceed; and it preserves LPL's structure and activity. Recently, GPIHBP1 was the key to solving the structure of LPL. These developments have transformed the models for intravascular triglyceride metabolism.
Keywords: chylomicronemia; endothelial cells; hypertriglyceridemia; lipid transport; lipoprotein lipase.
Copyright © 2019 Elsevier Inc. All rights reserved.
Figures

References
-
- Abifadel M, Jambart S, Allard D, Rabes JP, Varret M, Derre A, Chouery E, Salem N, Junien C, Aydenian H, et al. (2004). Identification of the first Lebanese mutation in the LPL gene and description of a rapid detection method. Clin. Genet 65, 158–161. - PubMed
-
- Acton S, Rigotti A, Landschulz KT, Xu S, Hobbs HH, and Krieger M (1996). Identification of scavenger receptor SR-BI as a high density lipoprotein receptor. Science 271, 518–520. - PubMed
