

The Cardiac Microenvironment Instructs Divergent Monocyte Fates and Functions in Myocarditis
- PMID: 31269438
- PMCID: PMC6813836
- DOI: 10.1016/j.celrep.2019.06.007
The Cardiac Microenvironment Instructs Divergent Monocyte Fates and Functions in Myocarditis
Abstract
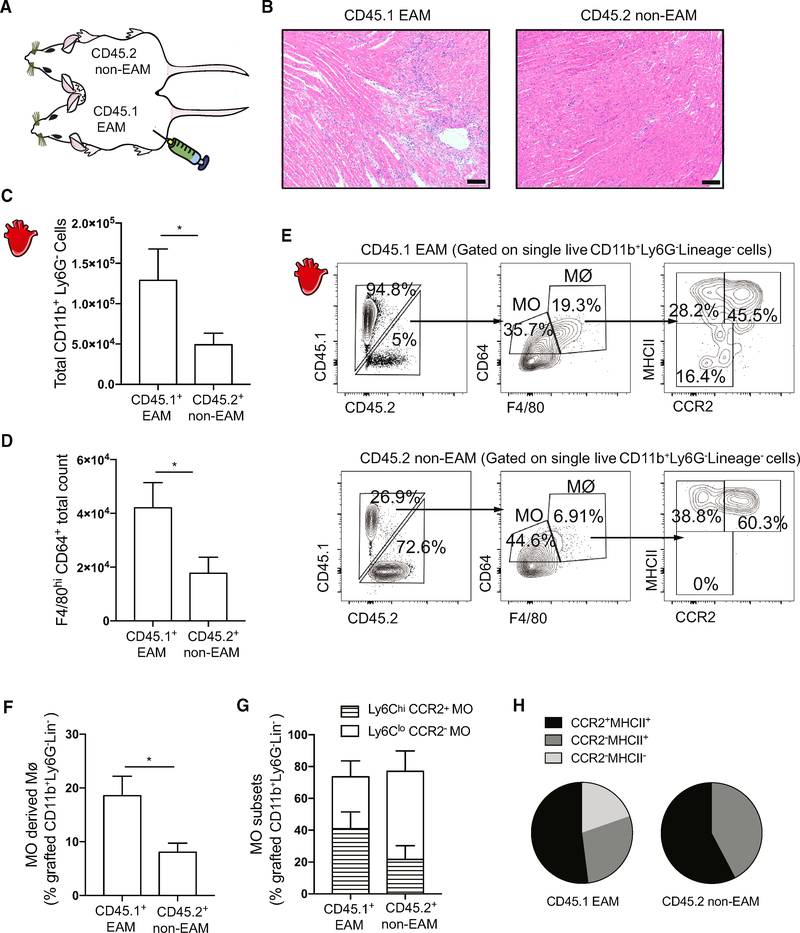
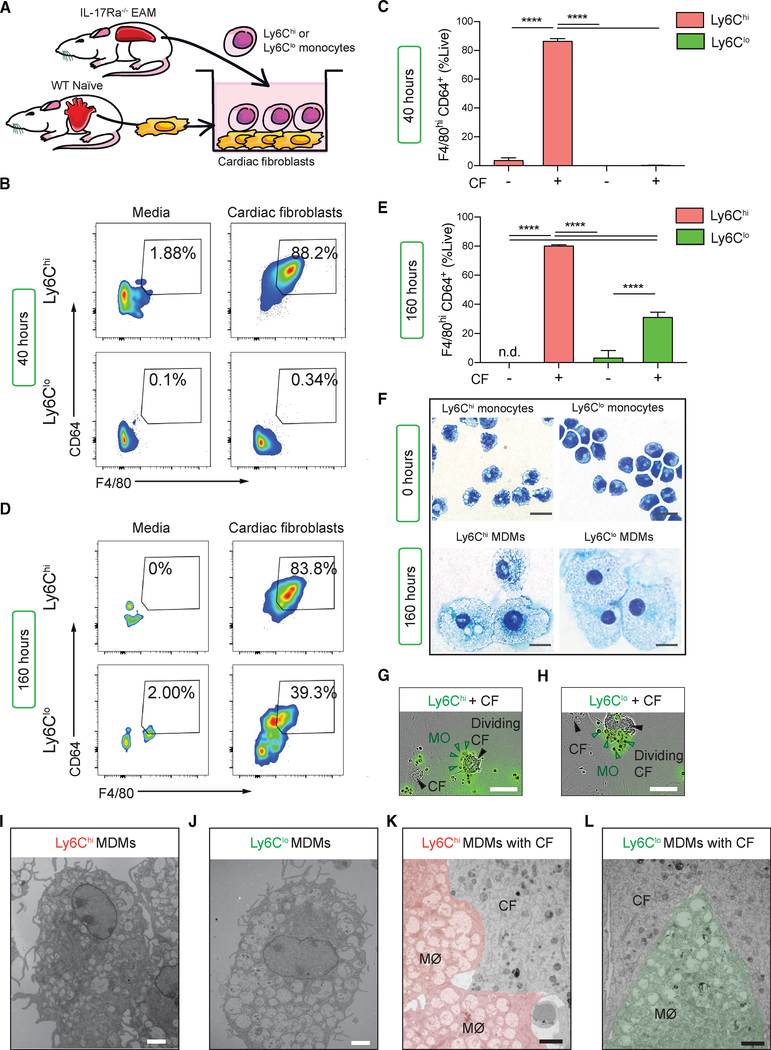
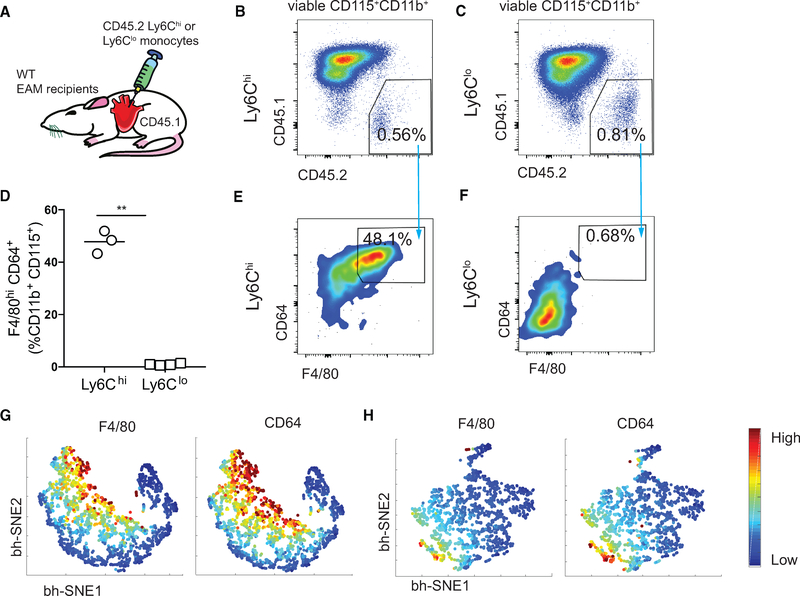
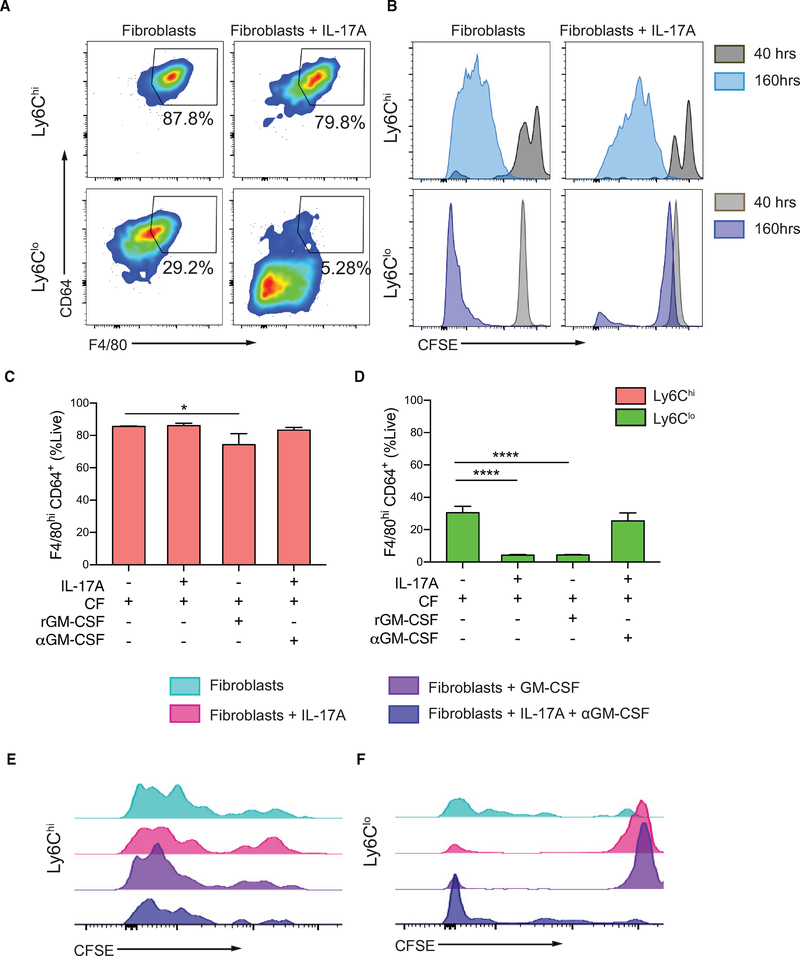
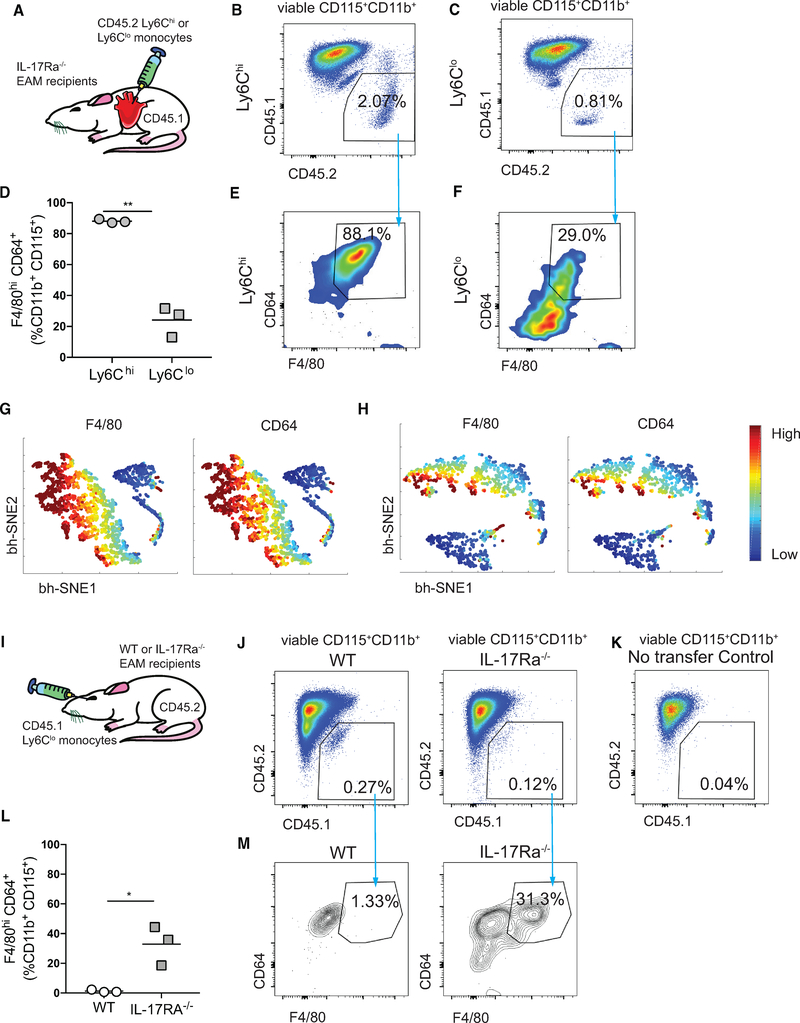
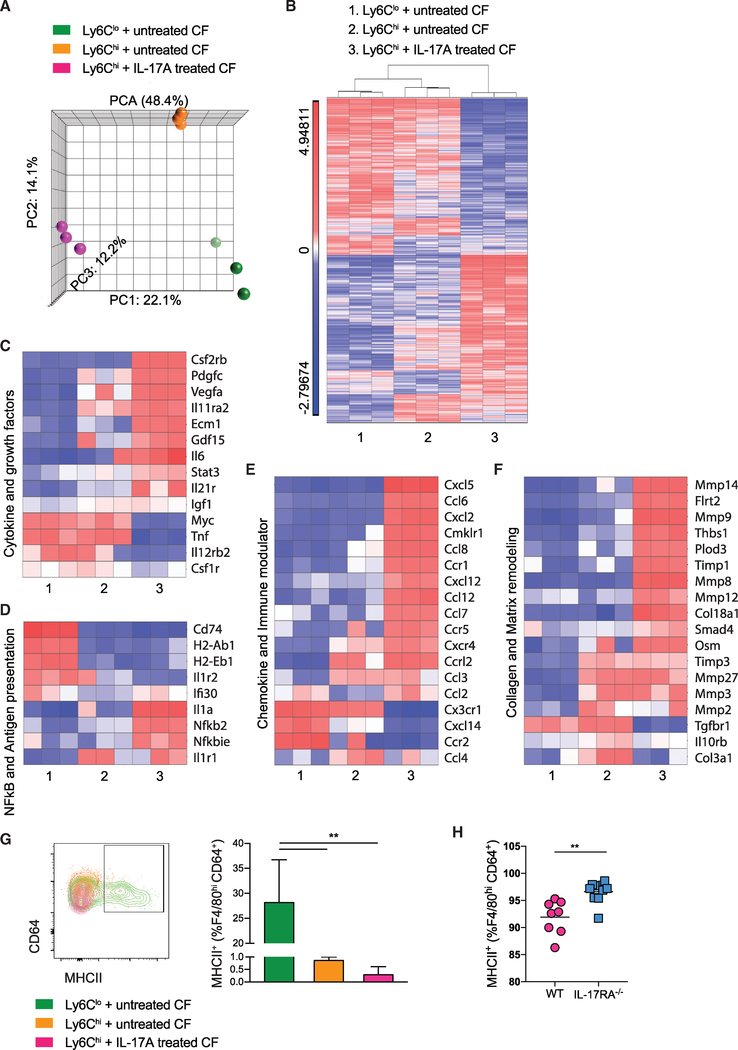
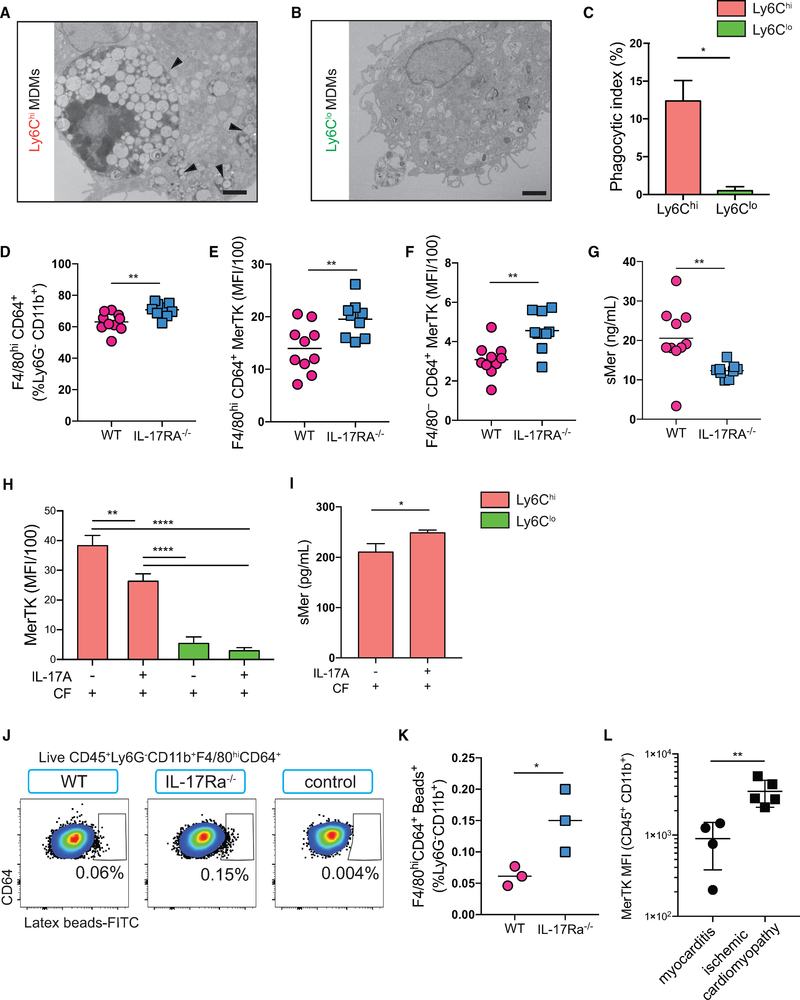
Two types of monocytes, Ly6Chi and Ly6Clo, infiltrate the heart in murine experimental autoimmune myocarditis (EAM). We discovered a role for cardiac fibroblasts in facilitating monocyte-to-macrophage differentiation of both Ly6Chi and Ly6Clo cells, allowing these macrophages to perform divergent functions in myocarditis progression. During the acute phase of EAM, IL-17A is highly abundant. It signals through cardiac fibroblasts to attenuate efferocytosis of Ly6Chi monocyte-derived macrophages (MDMs) and simultaneously prevents Ly6Clo monocyte-to-macrophage differentiation. We demonstrated an inverse clinical correlation between heart IL-17A levels and efferocytic receptor expressions in humans with heart failure (HF). In the absence of IL-17A signaling, Ly6Chi MDMs act as robust phagocytes and are less pro-inflammatory, whereas Ly6Clo monocytes resume their differentiation into MHCII+ macrophages. We propose that MHCII+Ly6Clo MDMs are associated with the reduction of cardiac fibrosis and prevention of the myocarditis sequalae.
Keywords: Ly6C; MerTK; heart; macrophages; monocytes; myocarditis.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
References
-
- Amit I, Winter DR, and Jung S (2016). The role of the local environment and epigenetics in shaping macrophage identity and their effect on tissue homeostasis. Nat. Immunol. 17, 18–25. - PubMed
-
- Amoah BP, Yang H, Zhang P, Su Z, and Xu H (2015). Immunopathogenesis of Myocarditis: The Interplay Between Cardiac Fibroblast Cells, Dendritic Cells, Macrophages and CD4+ T Cells. Scand. J. Immunol. 82, 1–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
