

Identification of the E3 Ligase TRIM29 as a Critical Checkpoint Regulator of NK Cell Functions
- PMID: 31270148
- PMCID: PMC6684831
- DOI: 10.4049/jimmunol.1900171
Identification of the E3 Ligase TRIM29 as a Critical Checkpoint Regulator of NK Cell Functions
Abstract
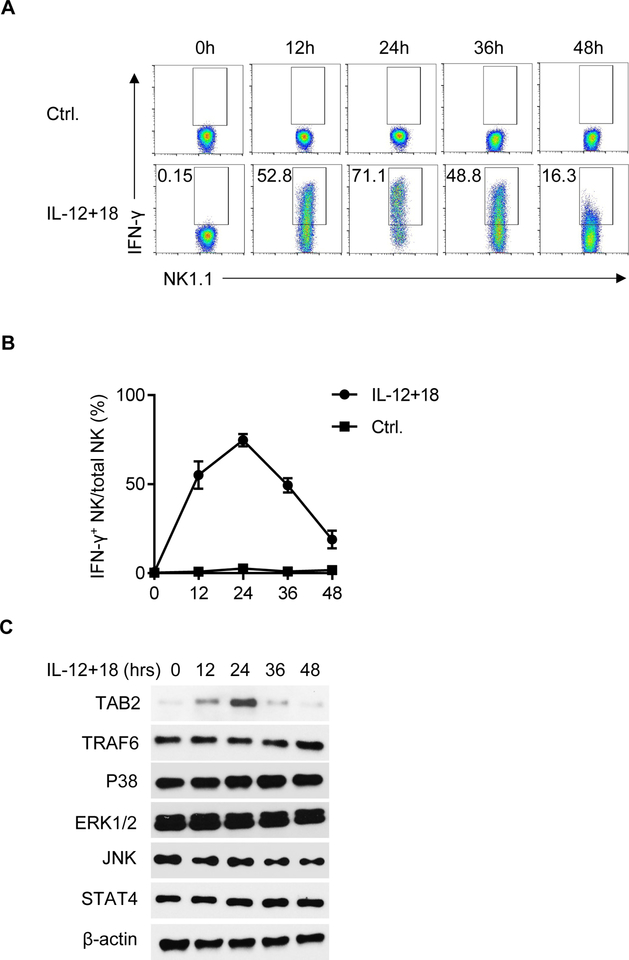
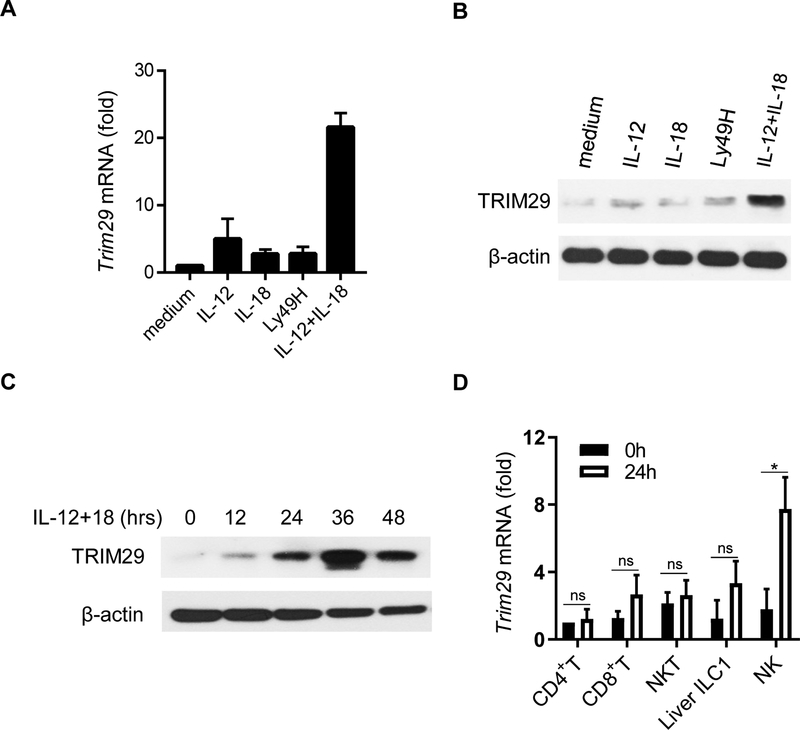
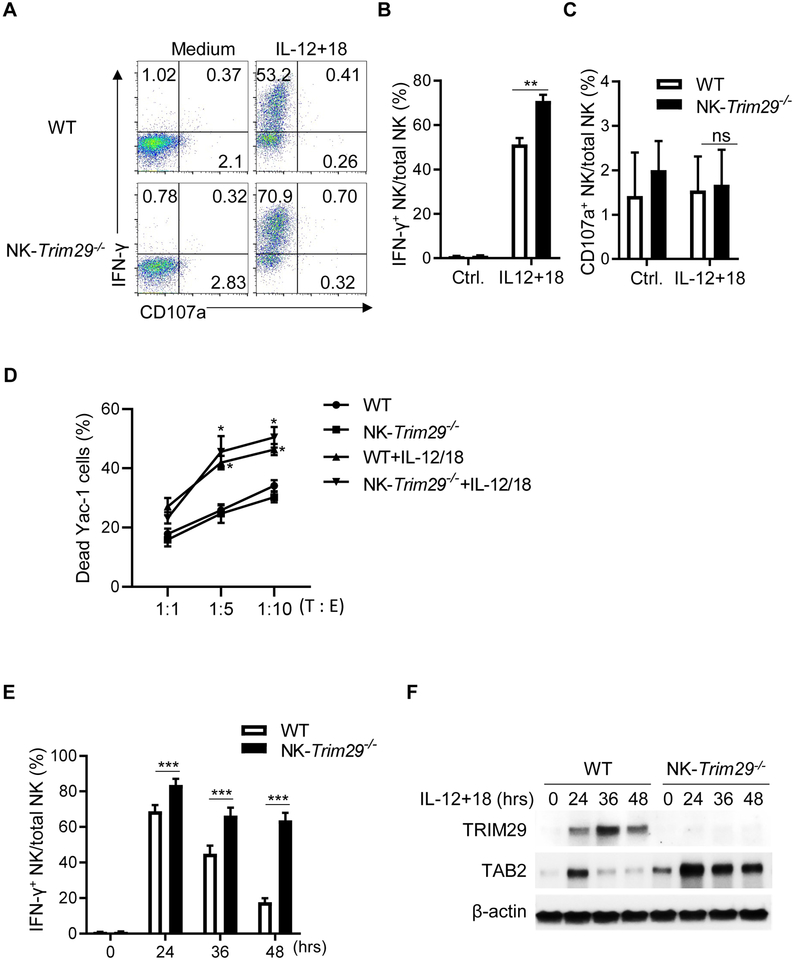
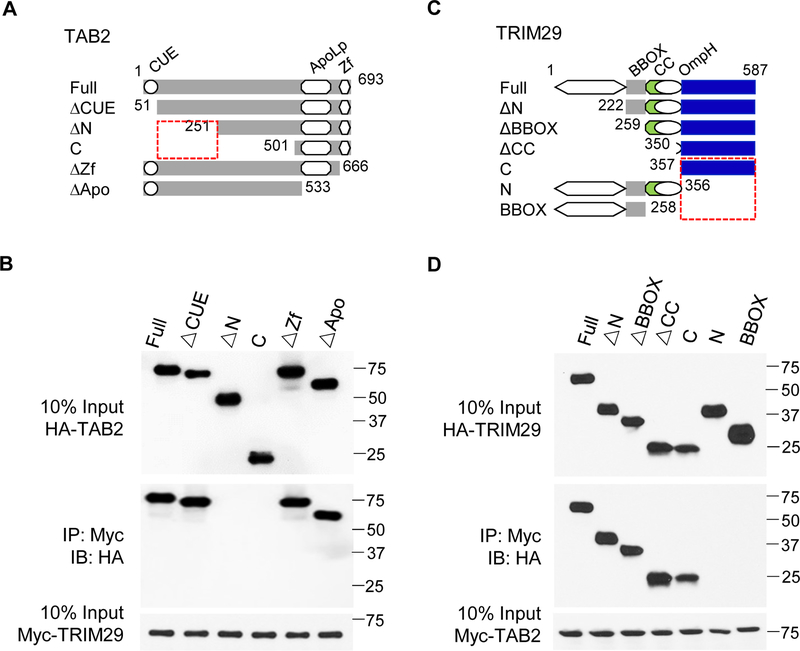
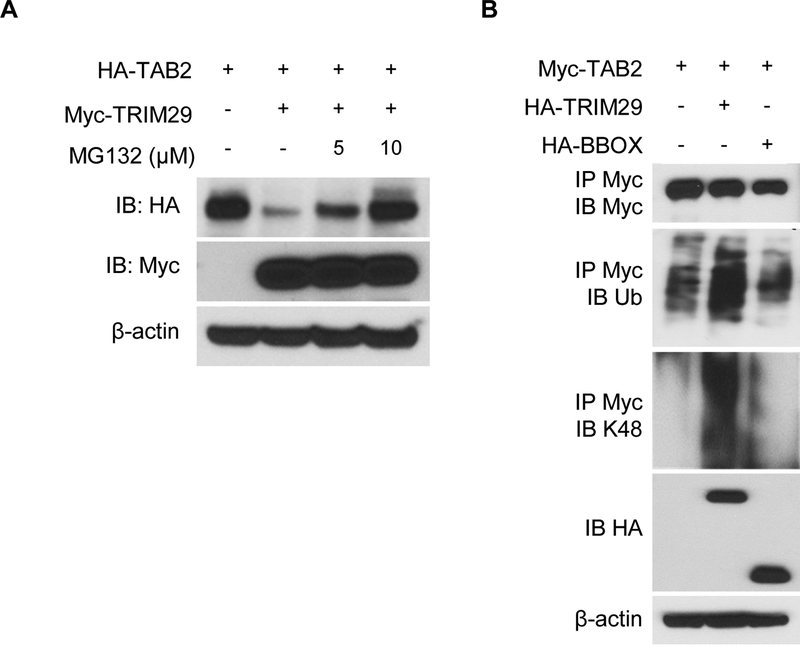
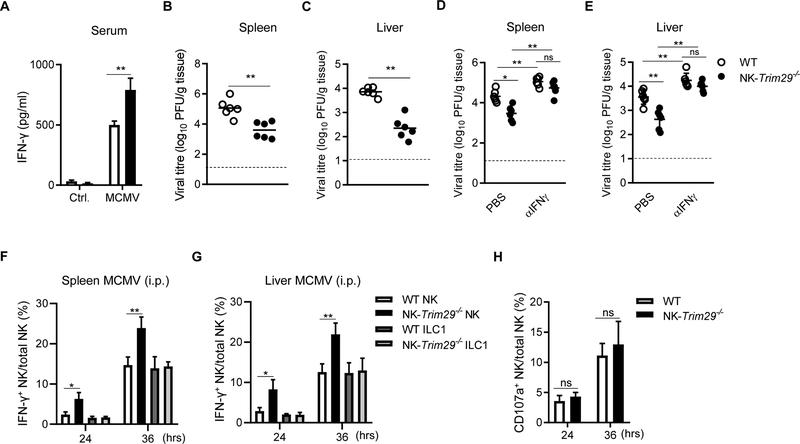
NK cells play an important role in immune surveillance and protective immunity, mainly through rapid cytokine release and cytolytic activities. But how such responses are negatively regulated remains poorly defined. In this study, we demonstrated that the E3 ubiquitin ligase TRIM29 is a crucial regulator of NK cell functions. We found that TRIM29 was not expressed in resting NK cells, but was readily upregulated following activation, especially after IL-12 plus IL-18 stimulation. The levels of TRIM29 expression were inversely correlated with IFN-γ production by NK cells, suggesting that TRIM29 inhibits NK cell functions. Indeed, deficiency of TRIM29, specifically in NK cells, resulted in an enhanced IFN-γ production and consequently protected mice from murine CMV infection. Mechanistically, we showed that once induced in NK cells, TRIM29 ubiquitinates and degrades the TGF-β-activated kinase 1 binding protein 2 (TAB2), a key adaptor protein in IFN-γ production by NK cells. These results identify TRIM29 as a negative regulator of NK cell functions and may have important clinical implications.
Copyright © 2019 by The American Association of Immunologists, Inc.
Conflict of interest statement
Disclosures
The authors declare no competing financial interests.
Figures
References
-
- Lanier LL 2005. NK cell recognition. Annu Rev Immunol 23: 225–274. - PubMed
-
- Chiossone L, Dumas PY, Vienne M, and Vivier E. 2018. Natural killer cells and other innate lymphoid cells in cancer. Nat Rev Immunol 18: 671–688. - PubMed
-
- Wensveen FM, Jelencic V, Valentic S, Sestan M, Wensveen TT, Theurich S, Glasner A, Mendrila D, Stimac D, Wunderlich FT, Bruning JC, Mandelboim O, and Polic B. 2015. NK cells link obesity-induced adipose stress to inflammation and insulin resistance. Nat Immunol 16: 376–385. - PubMed
-
- Gur C, Porgador A, Elboim M, Gazit R, Mizrahi S, Stern-Ginossar N, Achdout H, Ghadially H, Dor Y, Nir T, Doviner V, Hershkovitz O, Mendelson M, Naparstek Y, and Mandelboim O. 2010. The activating receptor NKp46 is essential for the development of type 1 diabetes. Nat Immunol 11: 121–128. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
