

Toxicity drives facilitation between 4 bacterial species
- PMID: 31270235
- PMCID: PMC6690002
- DOI: 10.1073/pnas.1906172116
Toxicity drives facilitation between 4 bacterial species
Abstract
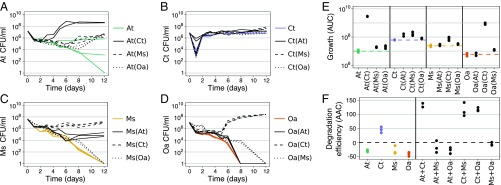
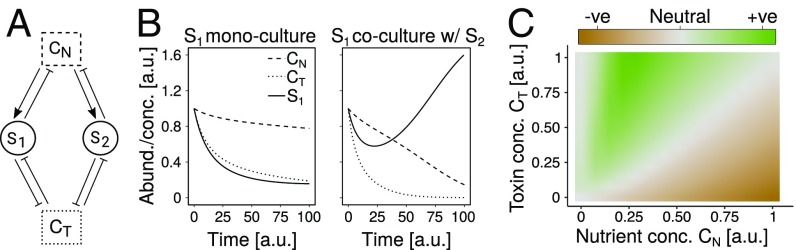
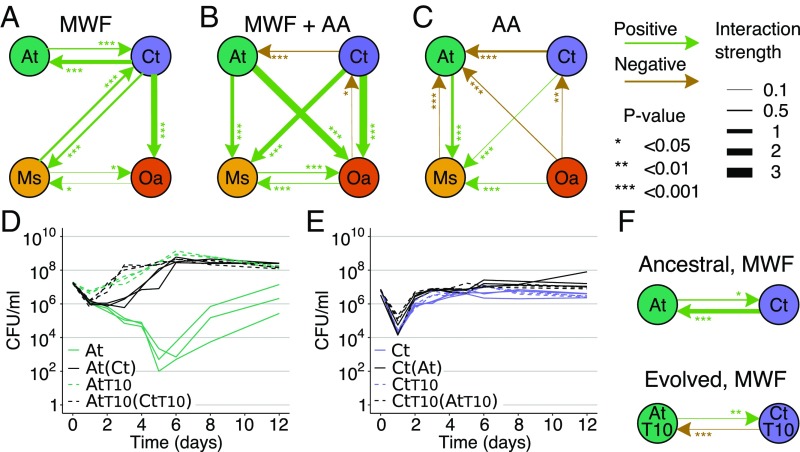
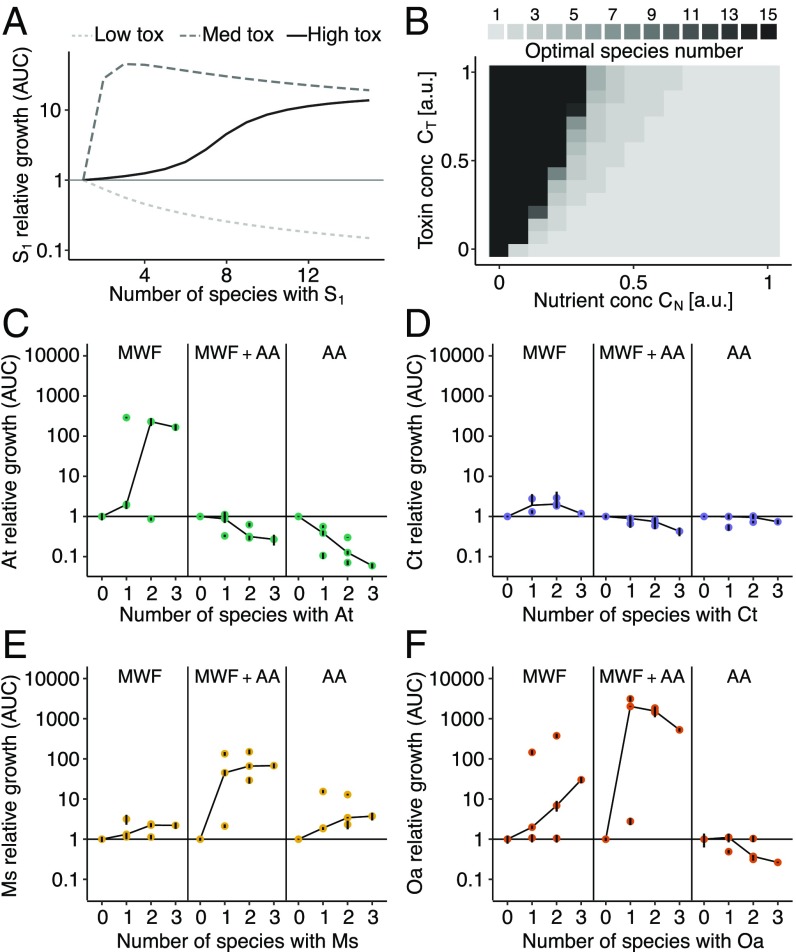
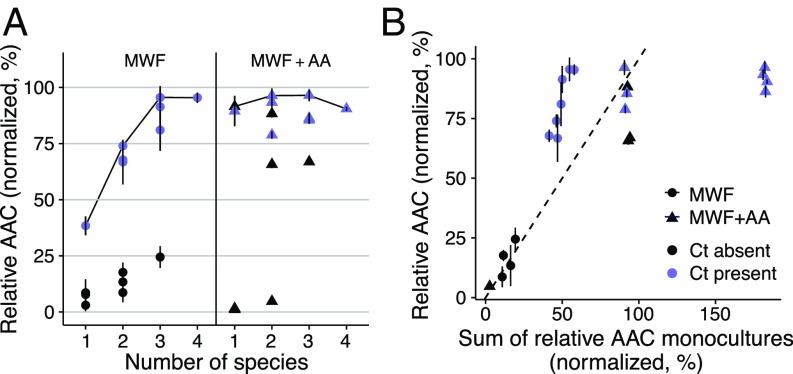
Competition between microbes is extremely common, with many investing in mechanisms to harm other strains and species. Yet positive interactions between species have also been documented. What makes species help or harm each other is currently unclear. Here, we studied the interactions between 4 bacterial species capable of degrading metal working fluids (MWF), an industrial coolant and lubricant, which contains growth substrates as well as toxic biocides. We were surprised to find only positive or neutral interactions between the 4 species. Using mathematical modeling and further experiments, we show that positive interactions in this community were likely due to the toxicity of MWF, whereby each species' detoxification benefited the others by facilitating their survival, such that they could grow and degrade MWF better when together. The addition of nutrients, the reduction of toxicity, or the addition of more species instead resulted in competitive behavior. Our work provides support to the stress gradient hypothesis by showing how harsh, toxic environments can strongly favor facilitation between microbial species and mask underlying competitive interactions.
Keywords: community function; competition; cooperation; species diversity; stress gradient hypothesis.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Refining the stress gradient hypothesis in a microbial community.Proc Natl Acad Sci U S A. 2019 Aug 6;116(32):15760-15762. doi: 10.1073/pnas.1910420116. Epub 2019 Jul 18. Proc Natl Acad Sci U S A. 2019. PMID: 31320585 Free PMC article. No abstract available.
References
-
- De Roy K., Marzorati M., Van den Abbeele P., Van de Wiele T., Boon N., Synthetic microbial ecosystems: An exciting tool to understand and apply microbial communities. Environ. Microbiol. 16, 1472–1481 (2014). - PubMed
-
- Dolinšek J., Goldschmidt F., Johnson D. R., Synthetic microbial ecology and the dynamic interplay between microbial genotypes. FEMS Microbiol. Rev. 40, 961–979 (2016). - PubMed
-
- Atashgahi S., et al. , Prospects for harnessing biocide resistance for bioremediation and detoxification. Science 360, 743–746 (2018). - PubMed
-
- West S. A., Griffin A. S., Gardner A. S., Diggle S. P., Social evolution theory for microorganisms. Nat. Rev. Microbiol. 4, 597–607 (2006). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
