

Iron-dependent histone 3 lysine 9 demethylation controls B cell proliferation and humoral immune responses
- PMID: 31270335
- PMCID: PMC6610088
- DOI: 10.1038/s41467-019-11002-5
Iron-dependent histone 3 lysine 9 demethylation controls B cell proliferation and humoral immune responses
Abstract
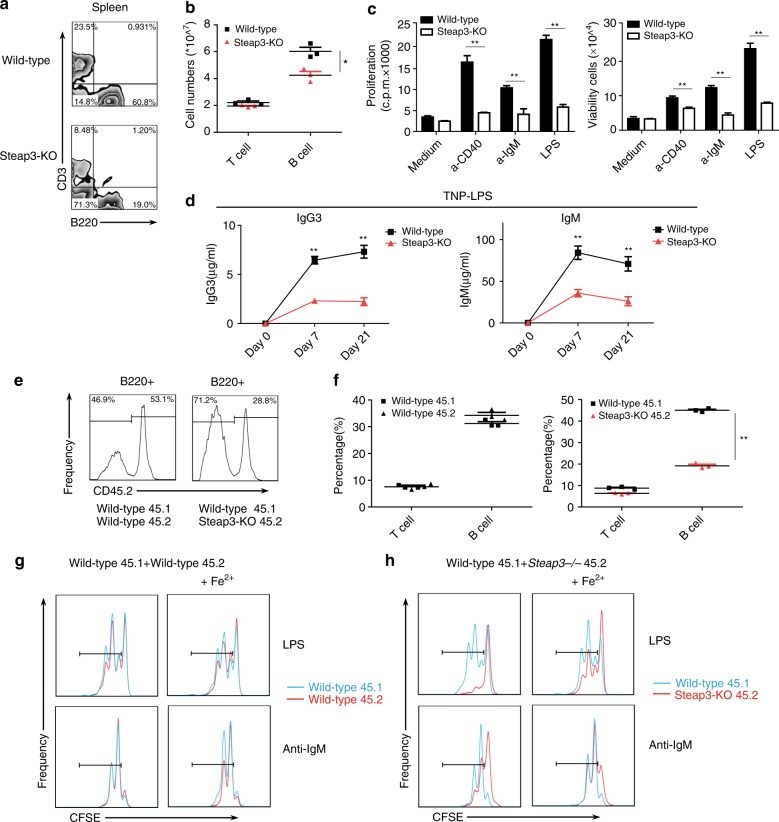
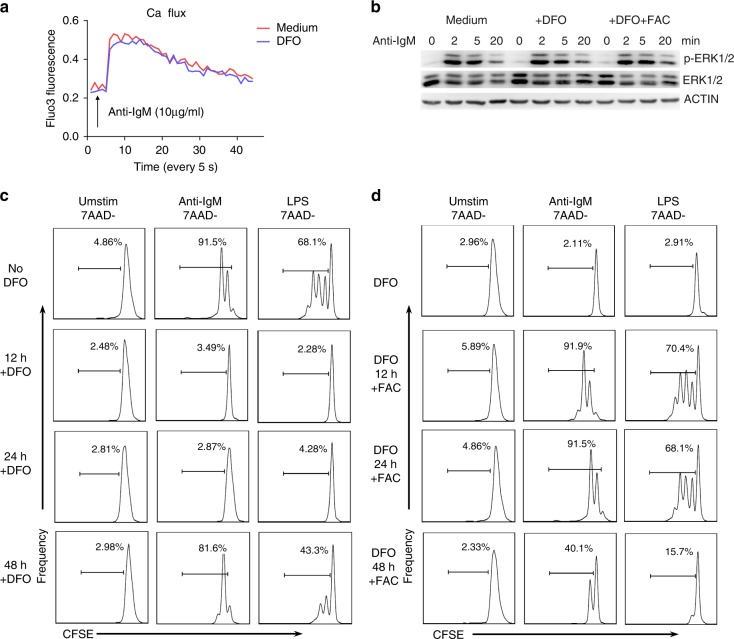
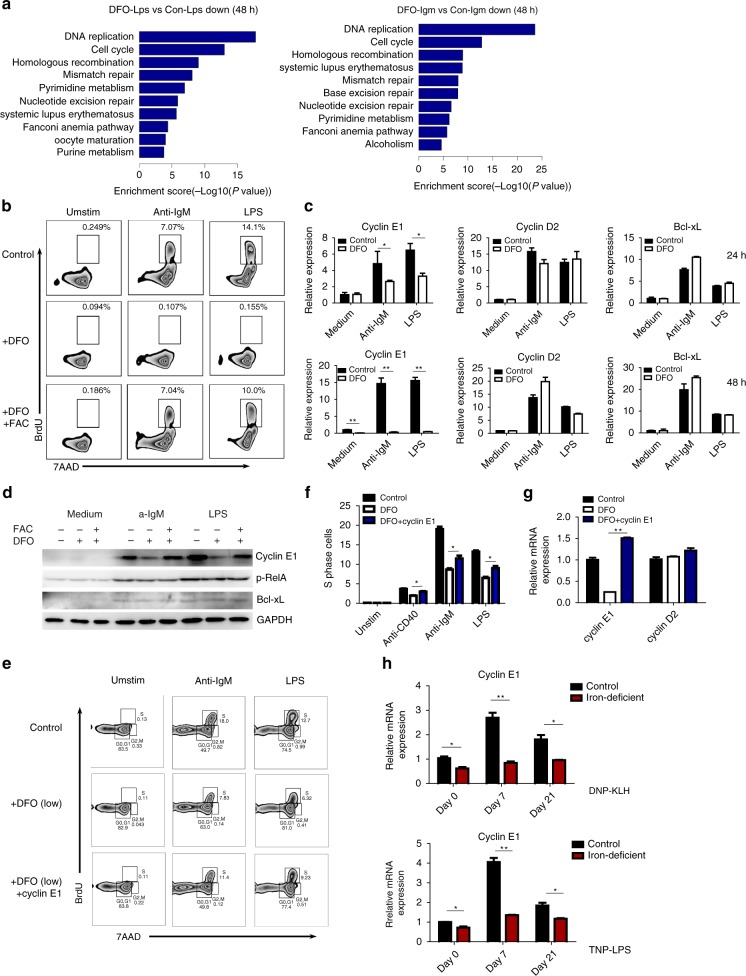
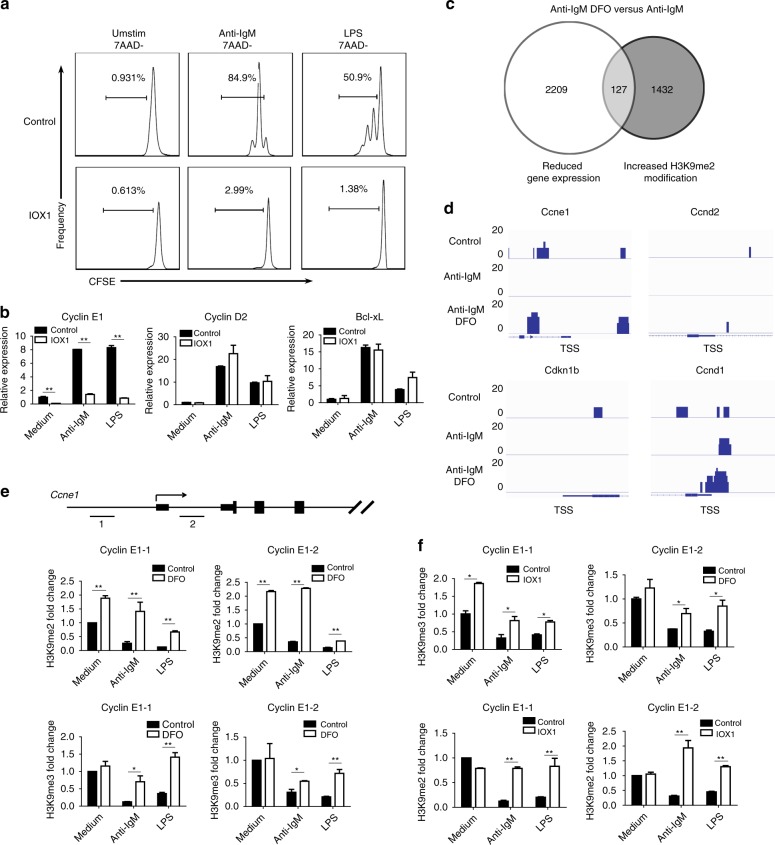
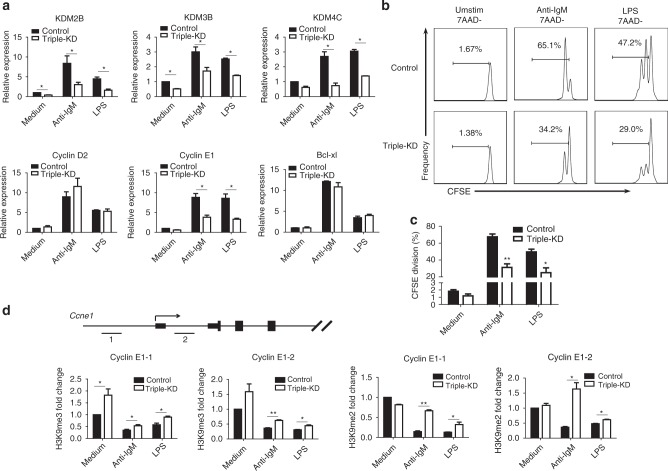
Trace elements play important roles in human health, but little is known about their functions in humoral immunity. Here, we show an important role for iron in inducing cyclin E and B cell proliferation. We find that iron-deficient individuals exhibit a significantly reduced antibody response to the measles vaccine when compared to iron-normal controls. Mice with iron deficiency also exhibit attenuated T-dependent or T-independent antigen-specific antibody responses. We show that iron is essential for B cell proliferation; both iron deficiency and α-ketoglutarate inhibition could suppress cyclin E1 induction and S phase entry of B cells upon activation. Finally, we demonstrate that three demethylases, KDM2B, KDM3B and KDM4C, are responsible for histone 3 lysine 9 (H3K9) demethylation at the cyclin E1 promoter, cyclin E1 induction and B cell proliferation. Thus, our data reveal a crucial role of H3K9 demethylation in B cell proliferation, and the importance of iron in humoral immunity.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
- 31530034/National Natural Science Foundation of China (National Science Foundation of China)/International
- 31570902/National Natural Science Foundation of China (National Science Foundation of China)/International
- 31370881/National Natural Science Foundation of China (National Science Foundation of China)/International
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
