

Identification of a novel EphB4 phosphodegron regulated by the autocrine IGFII/IRA axis in malignant mesothelioma
- PMID: 31270394
- PMCID: PMC8075896
- DOI: 10.1038/s41388-019-0854-y
Identification of a novel EphB4 phosphodegron regulated by the autocrine IGFII/IRA axis in malignant mesothelioma
Erratum in
-
Correction: Identification of a novel EphB4 phosphodegron regulated by the autocrine IGFII/IRA axis in malignant mesothelioma.Oncogene. 2021 Feb;40(8):1551. doi: 10.1038/s41388-020-01609-0. Oncogene. 2021. PMID: 33452463 Free PMC article. No abstract available.
Abstract
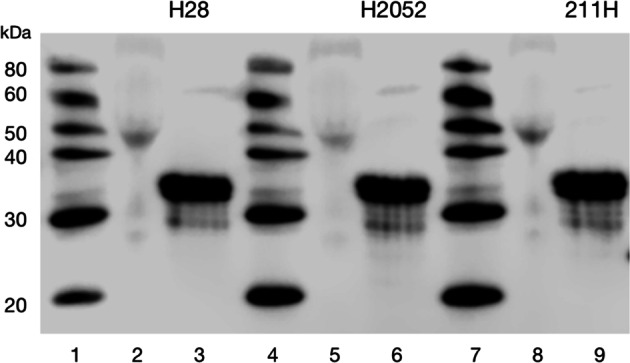
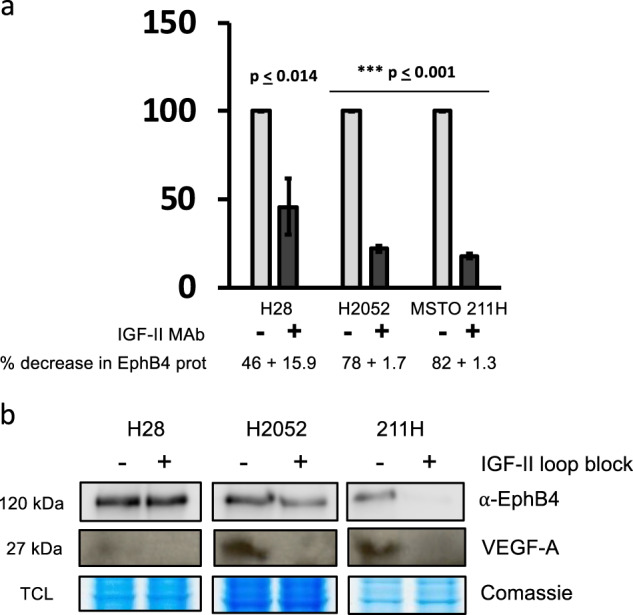
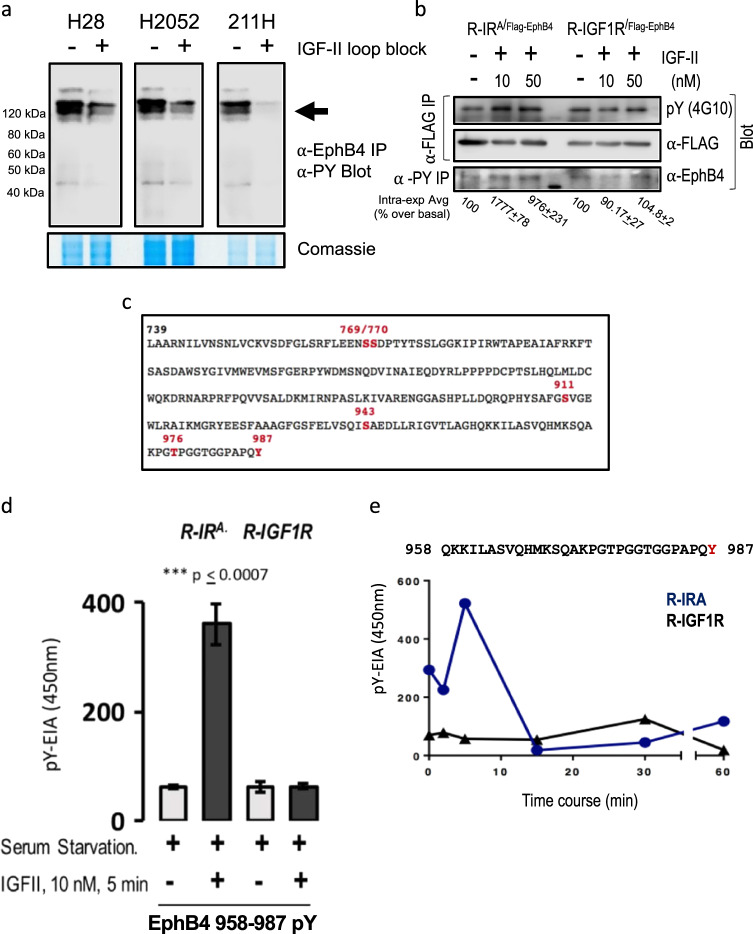
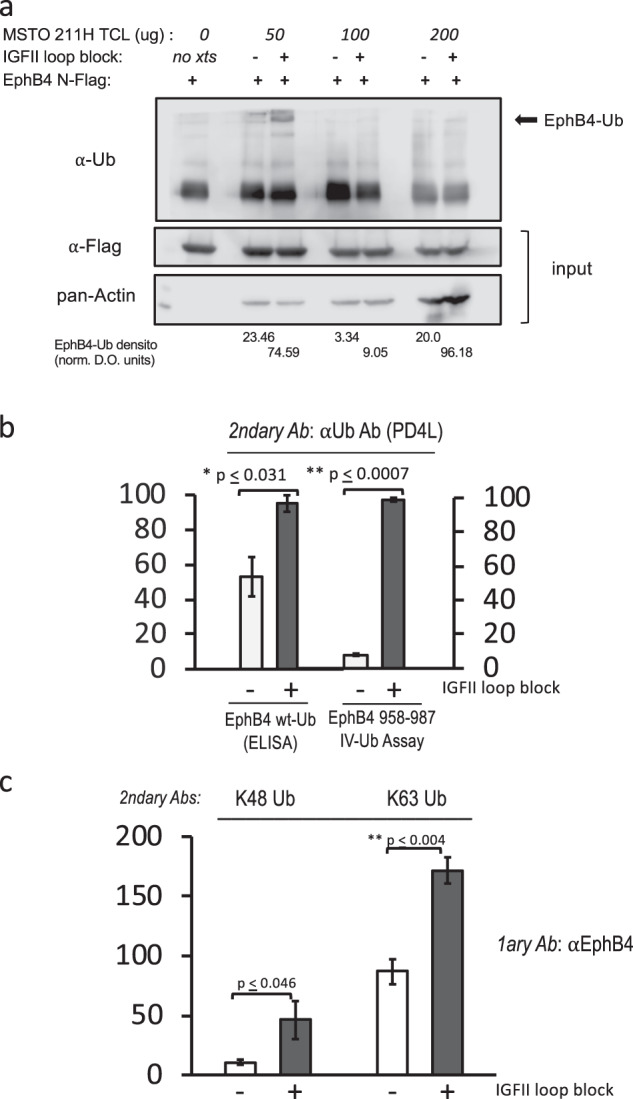
Malignant mesothelioma is a deadly disease with limited therapeutic options. EphB4 is an oncogenic tyrosine kinase receptor expressed in malignant mesothelioma as well as in a variety of cancers. It is involved in tumor microenvironment mediating angiogenesis and invasive cellular effects via both EphrinB2 ligand-dependent and independent mechanisms. The molecular network underlying EphB4 oncogenic effects is still unclear. Here we show that EphB4 expression in malignant mesothelioma cells is markedly decreased upon neutralization of cancer-secreted IGF-II. In particular, we demonstrate that EphB4 protein expression in malignant mesothelioma cells depend upon a degradation rescue mechanism controlled by the autocrine IGF-II-insulin receptor-A specific signaling axis. We show that the regulation of EphB4 expression is linked to a competing post-translational modification of its carboxy-terminal tail via phosphorylation of its tyrosine 987 by the Insulin receptor isoform-A kinase-associated activity in response to the autocrine IGF-II stimuli. Neutralization of this autocrine-induced EphB4-phosphorylation by IGF-II associates with the increased ubiquitination of EphB4 carboxy-terminal tail and with its rapid degradation. We also describe a novel Ubiquitin binding motif in the targeted region as part of the identified EphB4 phosphodegron and provide 3D modeling data supporting a possible model for the acute EphB4 PTM-driven regulation by IGF-II. Altogether, these findings disclose a novel molecular mechanism for the maintenance of EphB4-expression in malignant mesothelioma cells and other IGF-II-secreting cancers (IGF2omas).
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
-
- Wang HU, Chen ZF, Anderson DJ. Molecular distinction and angiogenic interaction between embryonic arteries and veins revealed by ephrin-B2 and its receptor Eph-B4. Cell. 1998;93:741–53. - PubMed
-
- Sinha UK, Kundra A, Scalia P, Smith DL, Parsa B, Masood R, et al. Expression of EphB4 in head and neck squamous cell carcinoma. Ear Nose Throat J. 2003;82:866. - PubMed
-
- Xia G, Kumar SR, Masood R, Zhu S, Reddy R, Krasnoperov V, et al. EphB4 expression and biological significance in prostate cancer. Cancer Res. 2005;65:4623–32. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
