

Molecular interactions underlying liquid-liquid phase separation of the FUS low-complexity domain
- PMID: 31270472
- PMCID: PMC6613800
- DOI: 10.1038/s41594-019-0250-x
Molecular interactions underlying liquid-liquid phase separation of the FUS low-complexity domain
Abstract
The low-complexity domain of the RNA-binding protein FUS (FUS LC) mediates liquid-liquid phase separation (LLPS), but the interactions between the repetitive SYGQ-rich sequence of FUS LC that stabilize the liquid phase are not known in detail. By combining NMR and Raman spectroscopy, mutagenesis, and molecular simulation, we demonstrate that heterogeneous interactions involving all residue types underlie LLPS of human FUS LC. We find no evidence that FUS LC adopts conformations with traditional secondary structure elements in the condensed phase; rather, it maintains conformational heterogeneity. We show that hydrogen bonding, π/sp2, and hydrophobic interactions all contribute to stabilizing LLPS of FUS LC. In addition to contributions from tyrosine residues, we find that glutamine residues also participate in contacts leading to LLPS of FUS LC. These results support a model in which FUS LC forms dynamic, multivalent interactions via multiple residue types and remains disordered in the densely packed liquid phase.
Conflict of interest statement
Competing Interests Statement
The authors declare no competing interests.
Figures
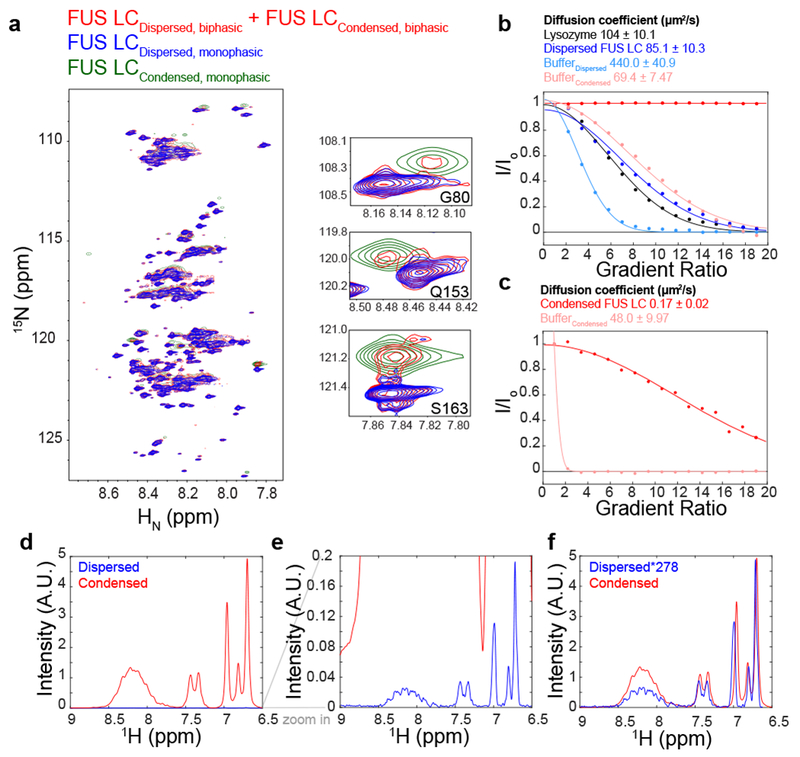
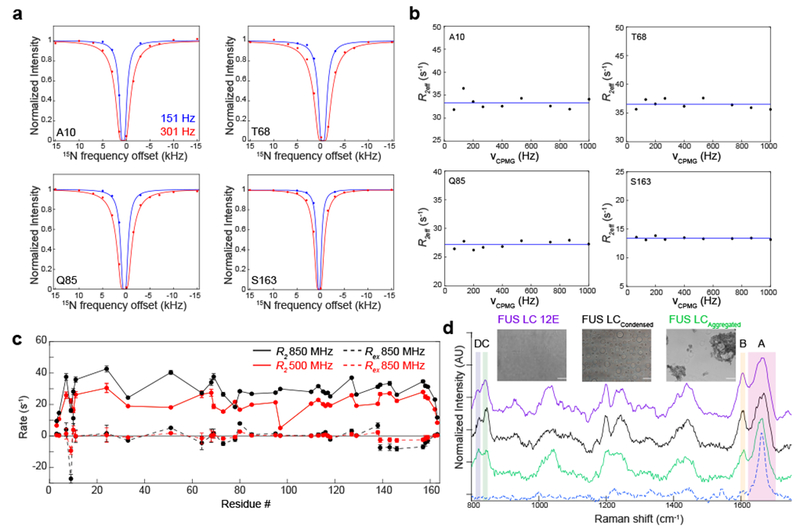
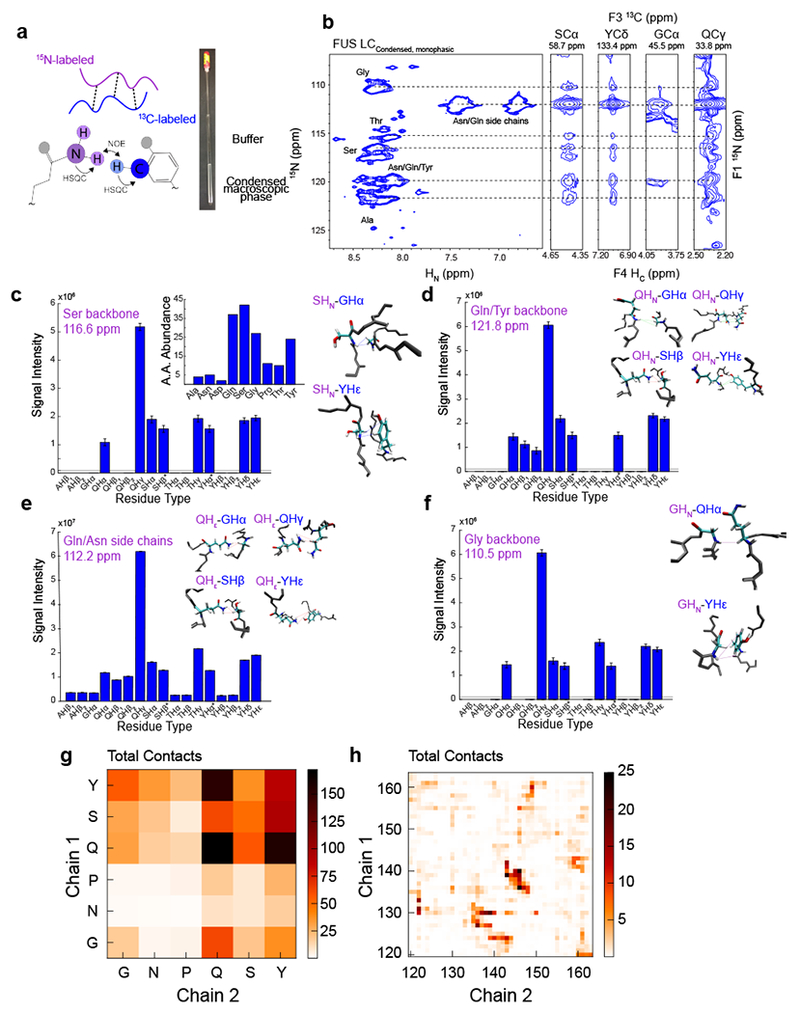
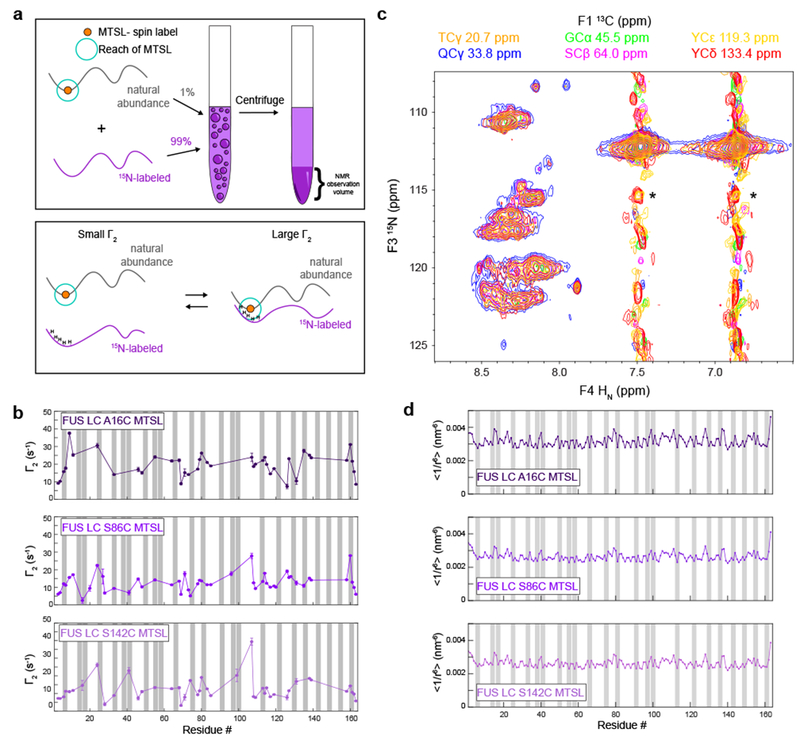
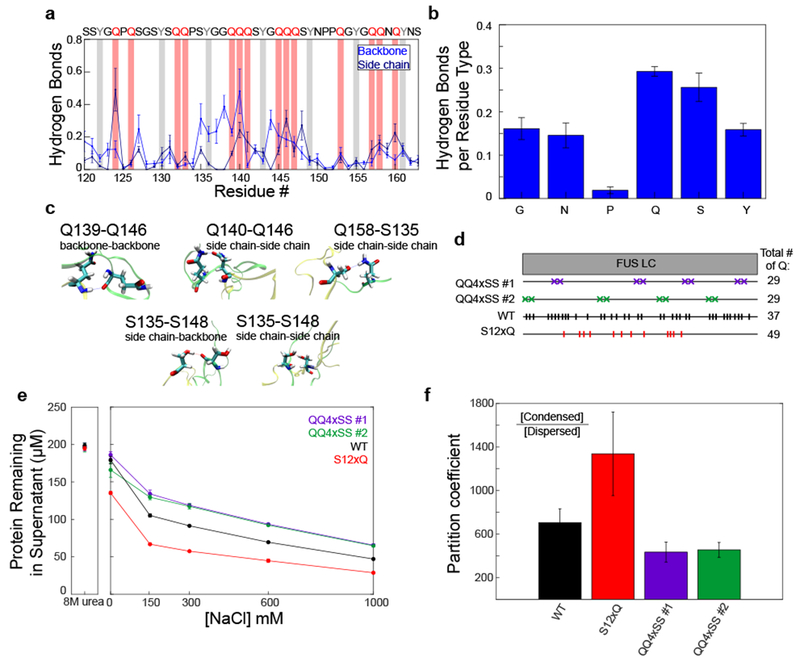
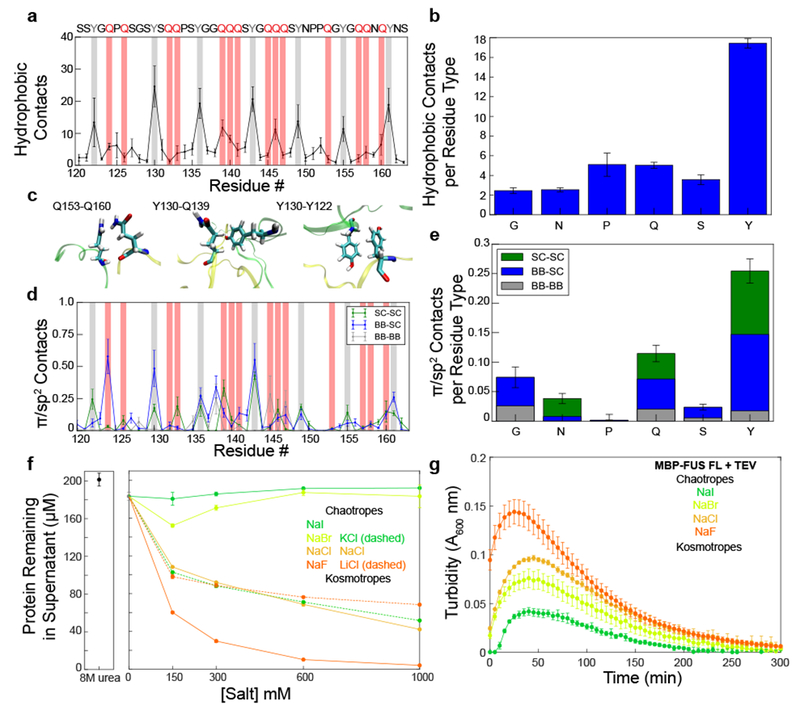
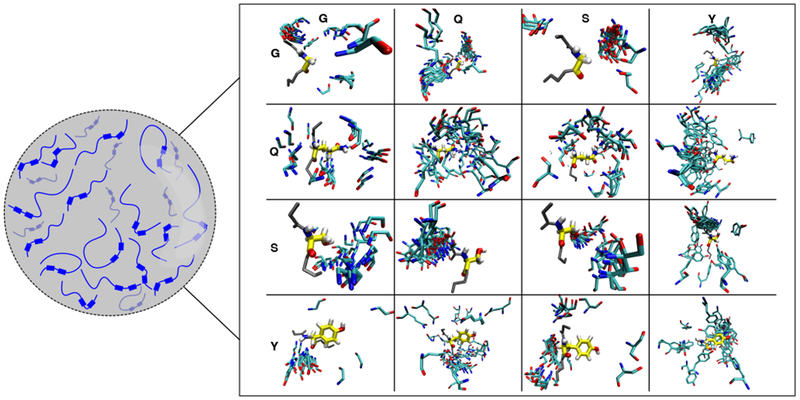
References
-
- Brangwynne CP et al. Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science (80-. ). 324, 1729–1732 (2009). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
