

Mechanics to pre-process information for the fine tuning of mechanoreceptors
- PMID: 31270587
- PMCID: PMC6726712
- DOI: 10.1007/s00359-019-01355-z
Mechanics to pre-process information for the fine tuning of mechanoreceptors
Abstract
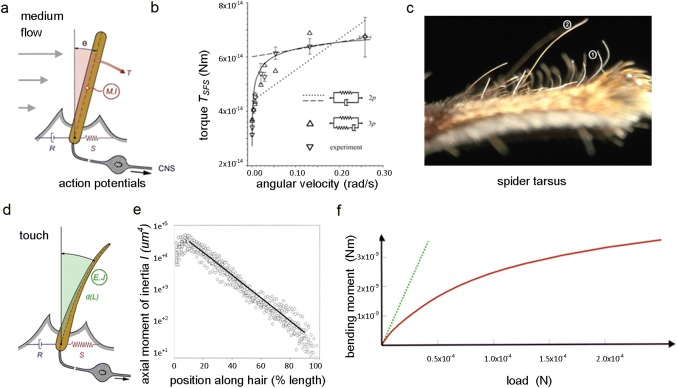
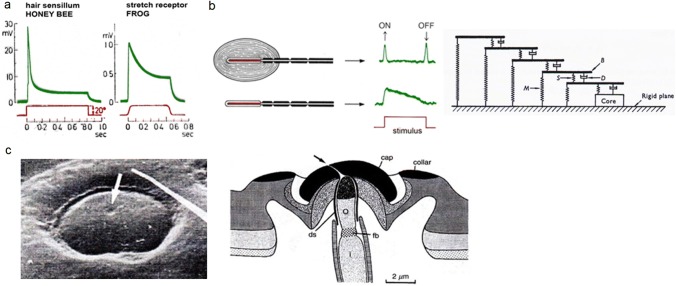
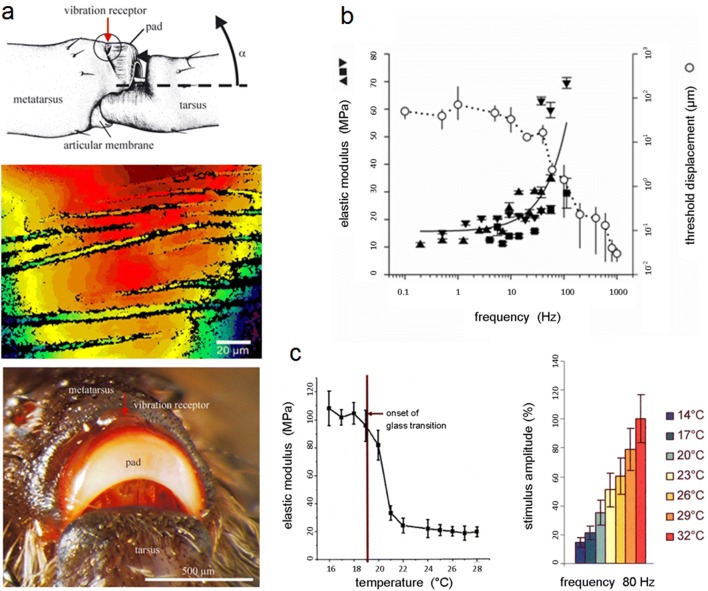
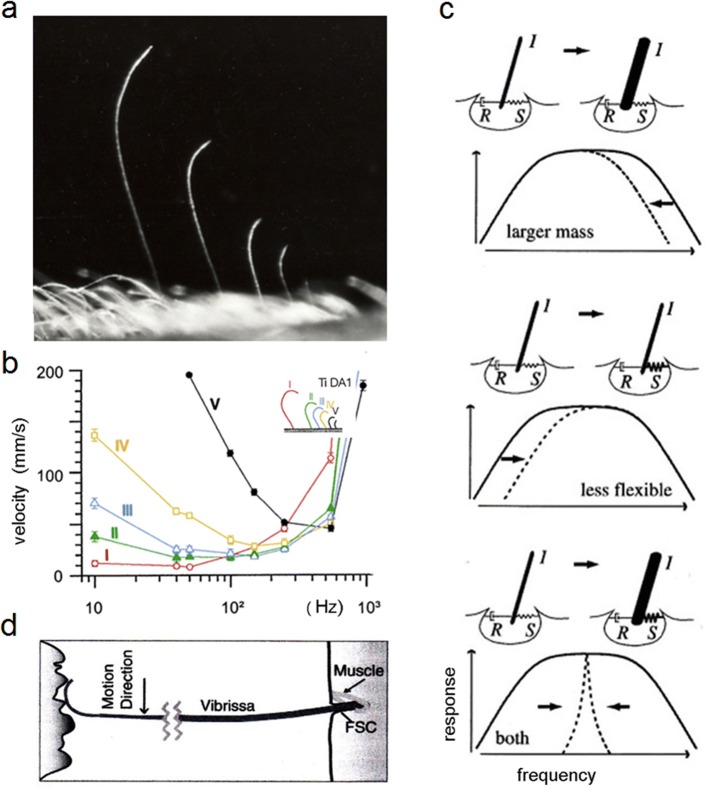
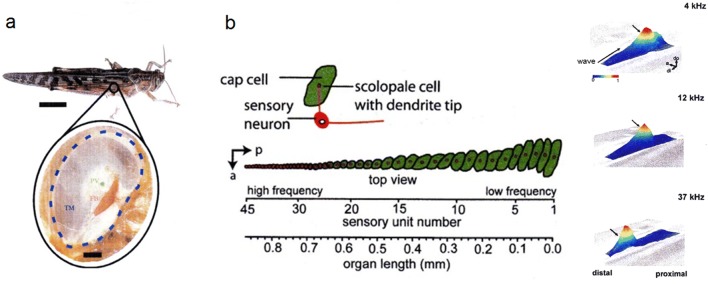
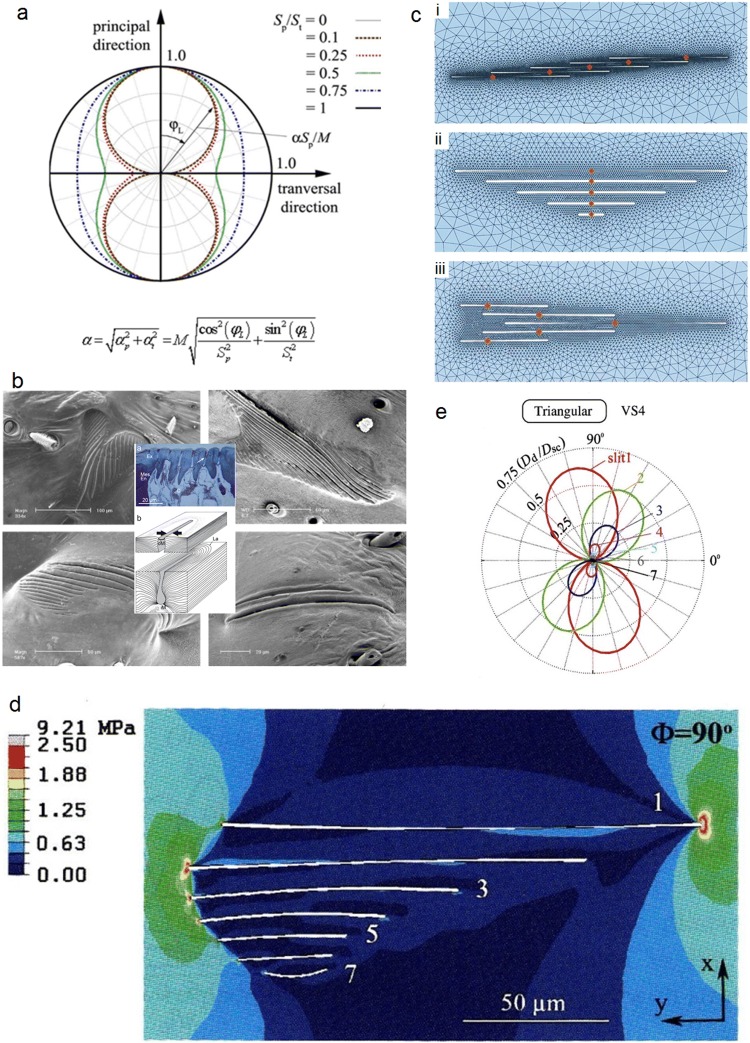
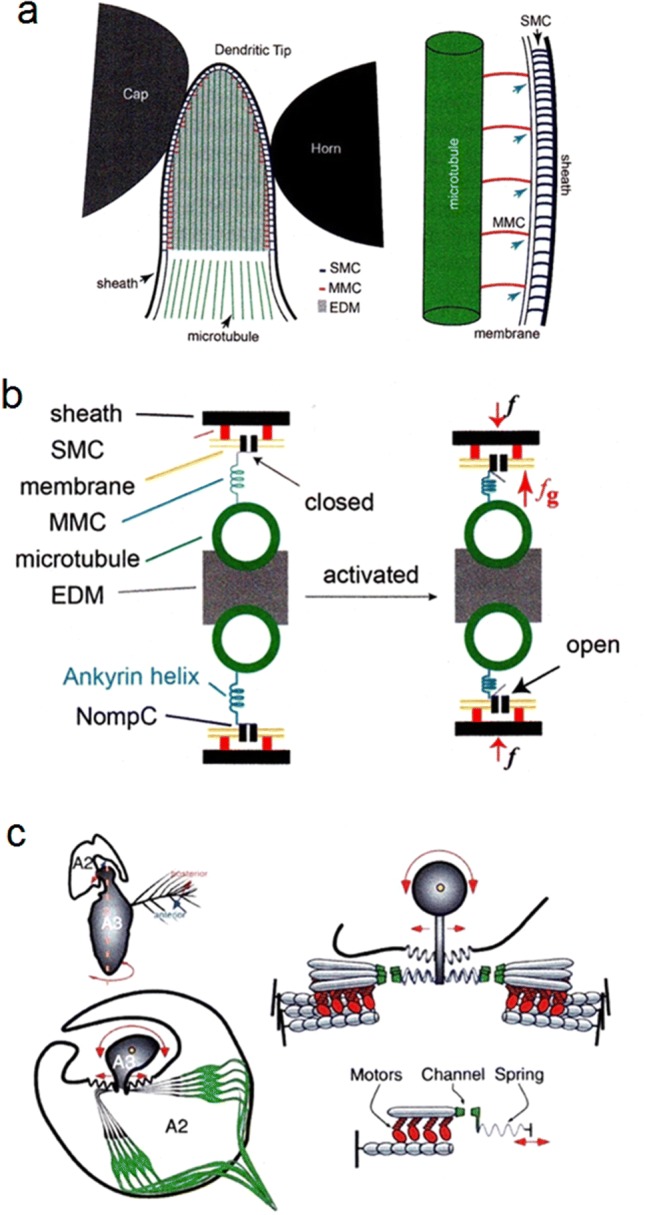
Non-nervous auxiliary structures play a significant role in sensory biology. They filter the stimulus and transform it in a way that fits the animal's needs, thereby contributing to the avoidance of the central nervous system's overload with meaningless stimuli and a corresponding processing task. The present review deals with mechanoreceptors mainly of invertebrates and some remarkable recent findings stressing the role of mechanics as an important source of sensor adaptedness, outstanding performance, and diversity. Instead of organizing the review along the types of stimulus energy (force) taken up by the sensors, processes associated with a few basic and seemingly simple mechanical principles like lever systems, viscoelasticity, resonance, traveling waves, and impedance matching are taken as the guideline. As will be seen, nature makes surprisingly competent use of such "simple mechanics".
Keywords: Auxiliary structures; Mechanoreception; Pre-processing of information; Sensor fine tuning; Stimulus transformation.
Figures
References
-
- Anton S, Barth FG. Central nervous projection patterns of trichobothria and other cuticular sensilla in the wandering spider Cupiennius salei (Arachnida, Araneae) Zoomorphol. 1993;113:21–32. doi: 10.1007/BF00430974. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
