

Human Corneal Expression of SLC4A11, a Gene Mutated in Endothelial Corneal Dystrophies
- PMID: 31273259
- PMCID: PMC6609610
- DOI: 10.1038/s41598-019-46094-y
Human Corneal Expression of SLC4A11, a Gene Mutated in Endothelial Corneal Dystrophies
Abstract
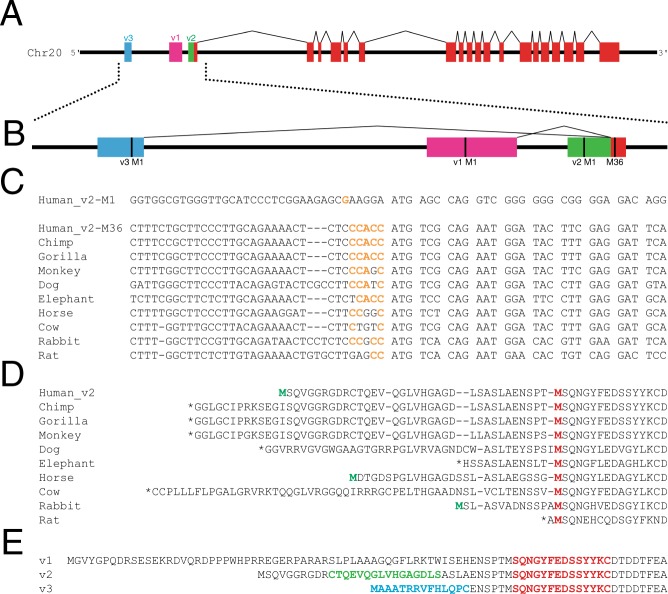
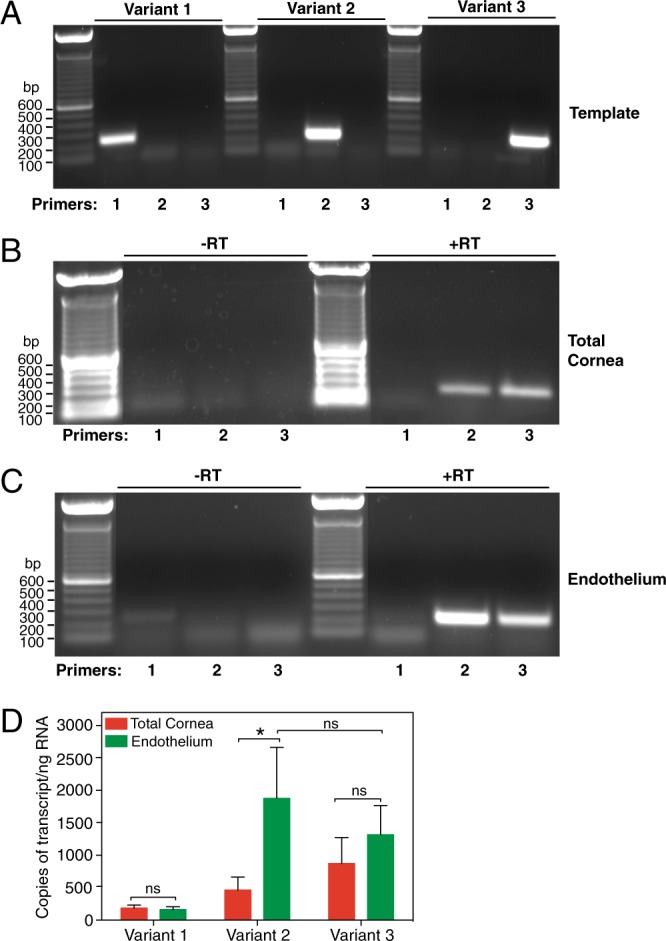
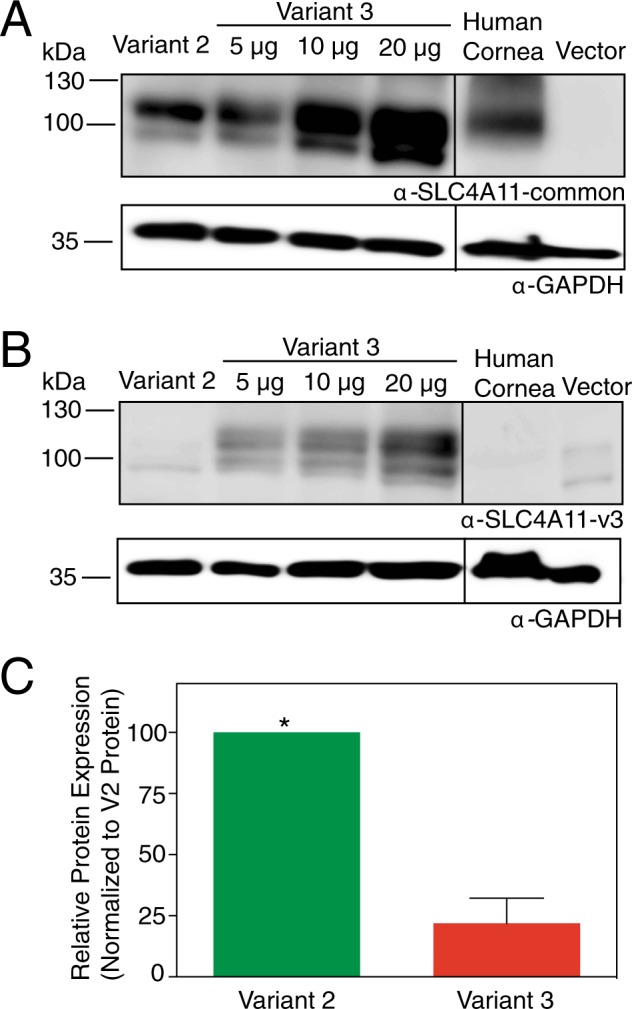
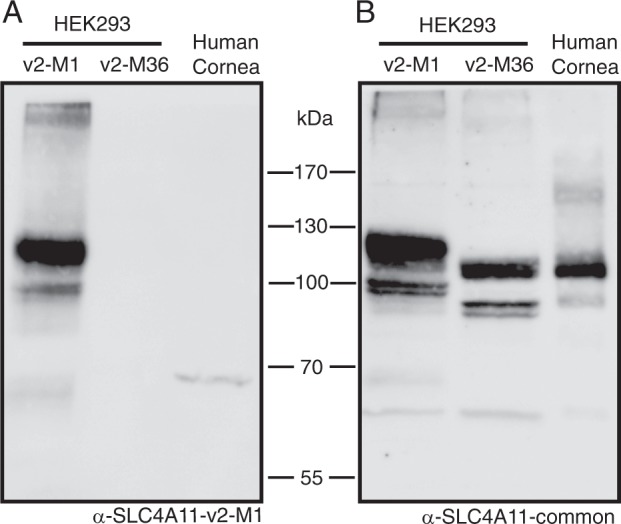
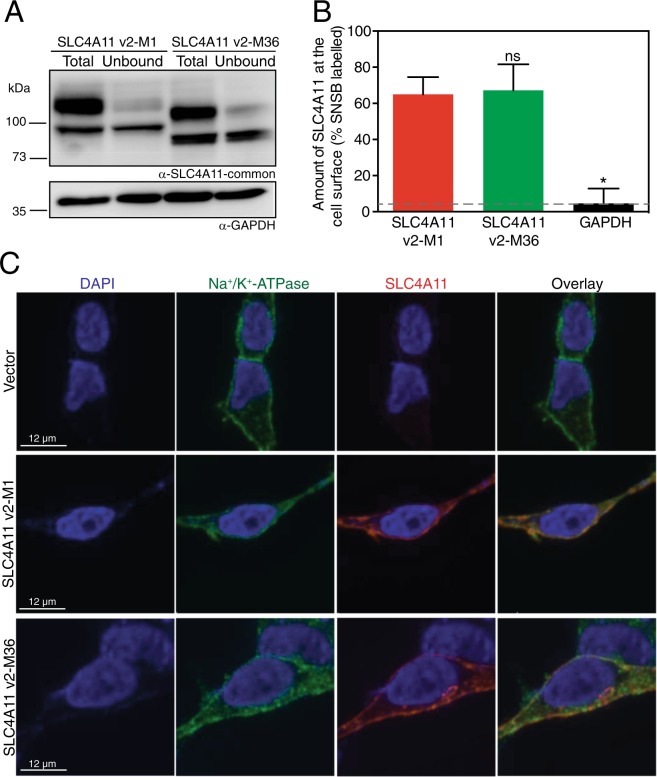
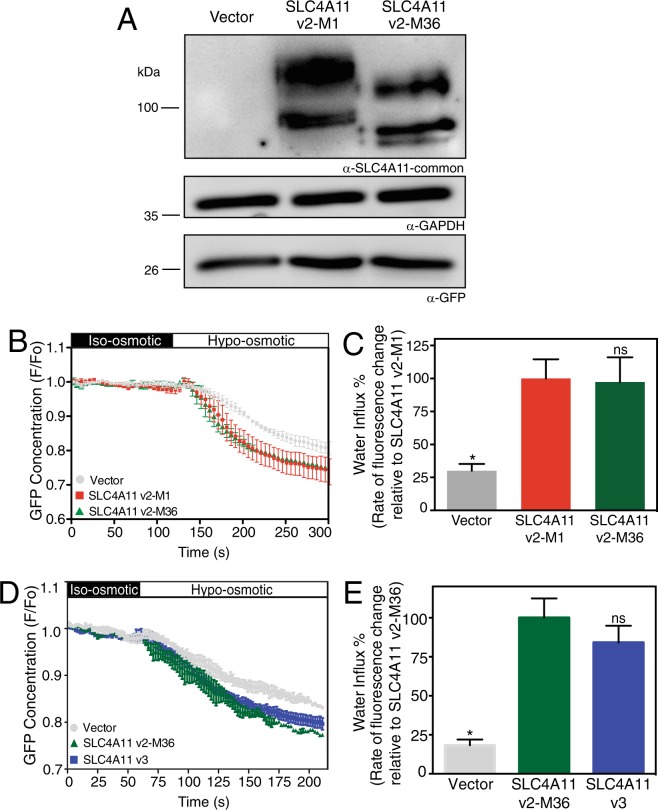
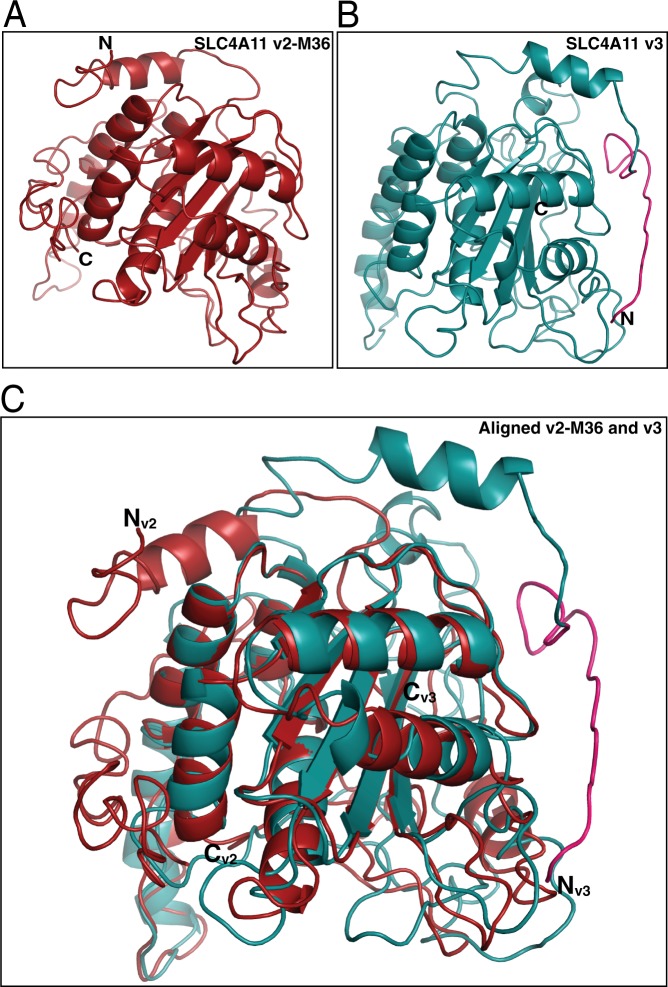
Two blinding corneal dystrophies, pediatric-onset congenital hereditary endothelial dystrophy (CHED) and some cases of late-onset Fuchs endothelial corneal dystrophy (FECD), are caused by SLC4A11 mutations. Three N-terminal SLC4A11 variants: v1, v2 and v3 are expressed in humans. We set out to determine which of these transcripts and what translated products, are present in corneal endothelium as these would be most relevant for CHED and FECD studies. Reverse transcription PCR (RT-PCR) and quantitative RT-PCR revealed only v2 and v3 mRNA in human cornea, but v2 was most abundant. Immunoblots probed with variant-specific antibodies revealed that v2 protein is about four times more abundant than v3 in human corneal endothelium. Bioinformatics and protein analysis using variant-specific antibodies revealed that second methionine in the open reading frame (M36) acts as translation initiation site on SLC4A11 v2 in human cornea. The v2 variants starting at M1 (v2-M1) and M36 (v2-M36) were indistinguishable in their cell surface trafficking and transport function (water flux). Structural homology models of v2-M36 and v3 suggest structural differences but their significance remains unclear. A combination of bioinformatics, RNA quantification and isoform-specific antibodies allows us to conclude that SLC4A11 variant 2 with start site M36 is predominant in corneal endothelium.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- 2016 Eye Banking Statisitical Report. (Eye Bank Association of America, Washington, D.C., U.S.A., 2017).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
