

A loss-of-function mutation in the DWARF4/ PETANKO5 gene enhances the late-flowering and semi-dwarf phenotypes of the Arabidopsis clock mutant lhy-12;cca1-101 under continuous light without affecting FLC expression
- PMID: 31274993
- PMCID: PMC6565938
- DOI: 10.5511/plantbiotechnology.16.0601a
A loss-of-function mutation in the DWARF4/ PETANKO5 gene enhances the late-flowering and semi-dwarf phenotypes of the Arabidopsis clock mutant lhy-12;cca1-101 under continuous light without affecting FLC expression
Abstract
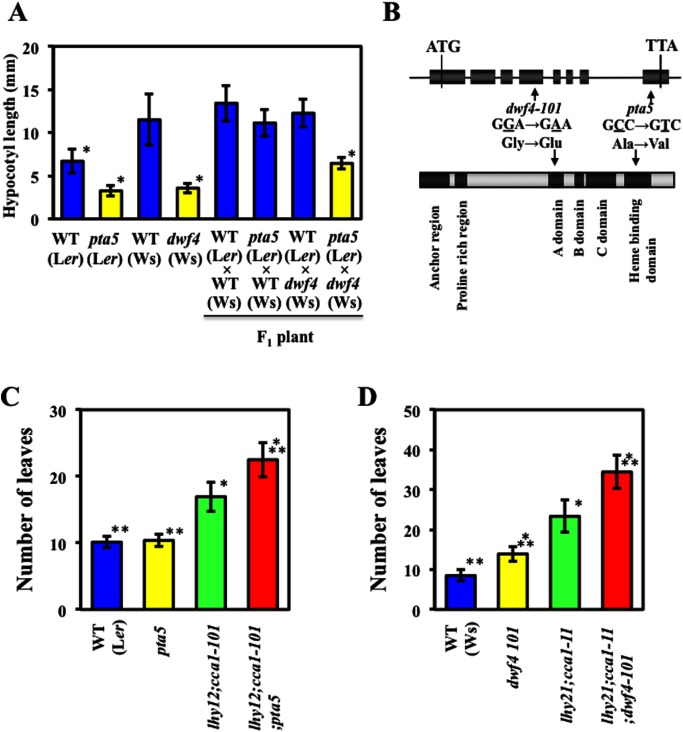
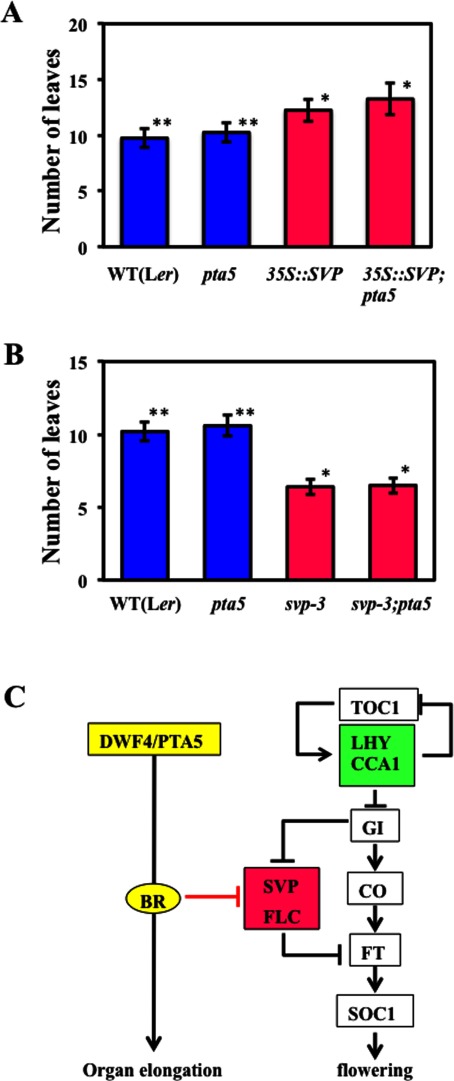
The circadian clock plays important roles in the control of photoperiodic flowering in Arabidopsis. Mutations in the LATE ELONGATED HYPOCOTYL (LHY) and CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) genes (lhy;cca1) accelerate flowering under short days, whereas lhy;cca1 delays flowering under continuous light (LL). The lhy;cca1 mutant also exhibits short hypocotyls and petioles under LL. However, the molecular mechanisms underlying the regulation of both flowering time and organ lengths in the LHY/CCA1-dependent pathway are not fully understood. To address these questions, we performed EMS mutagenesis of the lhy-12;cca1-101 line and screened for mutations that enhance the lhy;cca1 phenotypes under LL. In this screen, we identified a novel allele of dwarf4 (dwf4) and named it petanko 5 (pta5). A similar level of enhancement of the delay in flowering was observed in these two dwf4 mutants when combined with the lhy;cca1 mutations. The lhy;cca1 and dwf4 mutations did not significantly affect the expression level of the floral repressor gene FLC under LL. Our results suggest that a defect in brassinosteroid (BR) signaling delayed flowering independent of the FLC expression level, at least in plants with the lhy;cca1 mutation grown under LL. The dwf4/pta5 mutation did not enhance the late-flowering phenotype of plants overexpressing SVP under LL, suggesting that SVP and BR function in a common pathway that controls flowering time. Our results suggest that the lhy;cca1 mutant exhibits delayed flowering due to both the BR signaling-dependent and -independent pathways under LL.
Keywords: CCA1; DWF4; LHY; brassinosteroid; flowering time.
Figures


Similar articles
-
Suppression of late-flowering and semi-dwarf phenotypes in the Arabidopsis clock mutant lhy-12;cca1-101 by phyB under continuous light.Plant Signal Behav. 2011 Aug;6(8):1162-71. doi: 10.4161/psb.6.8.16361. Epub 2011 Aug 1. Plant Signal Behav. 2011. PMID: 21822060 Free PMC article.
-
Double loss-of-function mutation in EARLY FLOWERING 3 and CRYPTOCHROME 2 genes delays flowering under continuous light but accelerates it under long days and short days: an important role for Arabidopsis CRY2 to accelerate flowering time in continuous light.J Exp Bot. 2011 May;62(8):2731-44. doi: 10.1093/jxb/erq450. Epub 2011 Feb 4. J Exp Bot. 2011. PMID: 21296763
-
Possible role of early flowering 3 (ELF3) in clock-dependent floral regulation by short vegetative phase (SVP) in Arabidopsis thaliana.New Phytol. 2009 Jun;182(4):838-850. doi: 10.1111/j.1469-8137.2009.02809.x. New Phytol. 2009. PMID: 19383102
-
Shedding light on the circadian clock and the photoperiodic control of flowering.Curr Opin Plant Biol. 2003 Feb;6(1):13-9. doi: 10.1016/s1369-5266(02)00011-0. Curr Opin Plant Biol. 2003. PMID: 12495746 Review.
-
MYB transcription factors in the Arabidopsis circadian clock.J Exp Bot. 2002 Jul;53(374):1551-7. doi: 10.1093/jxb/erf027. J Exp Bot. 2002. PMID: 12096093 Review.
Cited by
-
Density effects on semi-dwarf and early flowering mutants of Arabidopsis thaliana under continuous light.Plant Biotechnol (Tokyo). 2016;33(4):333-339. doi: 10.5511/plantbiotechnology.16.0912a. Epub 2016 Nov 9. Plant Biotechnol (Tokyo). 2016. PMID: 31274995 Free PMC article.
-
Comparative Transcriptome Analysis of Genes Involved in Anthocyanin Biosynthesis in Red and Green Walnut (Juglans regia L.).Molecules. 2017 Dec 22;23(1):25. doi: 10.3390/molecules23010025. Molecules. 2017. PMID: 29271948 Free PMC article.
References
-
- Busov VB, Brunner AM, Strauss SH (2008) Genes for control of plant stature and form. New Phytol 177: 589–607 - PubMed
-
- Domagalska MA, Schomburg FM, Amasino RM, Vierstra RD, Nagy F, Davis SJ (2007) Attenuation of brassinosteroid signaling enhances FLC expression and delays flowering. Development 134: 2841–2850 - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases