

Chloroplast Protein Degradation in Senescing Leaves: Proteases and Lytic Compartments
- PMID: 31275332
- PMCID: PMC6593067
- DOI: 10.3389/fpls.2019.00747
Chloroplast Protein Degradation in Senescing Leaves: Proteases and Lytic Compartments
Abstract
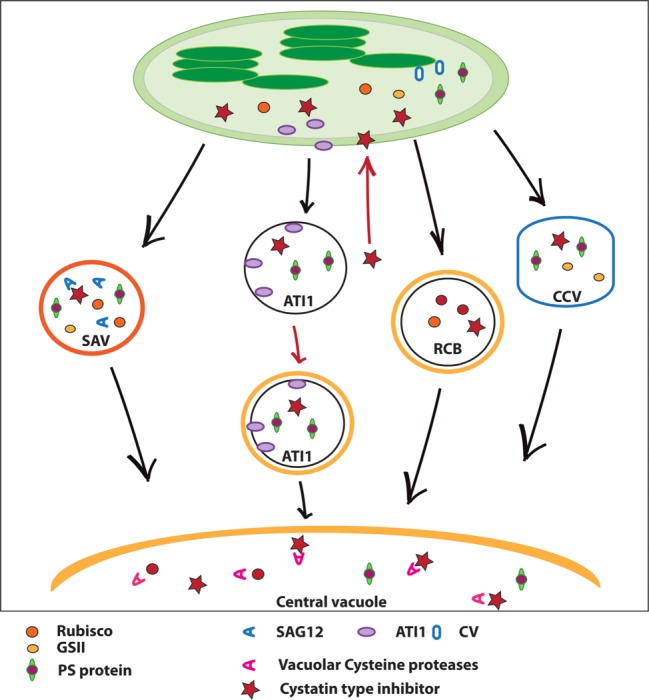
Leaf senescence is characterized by massive degradation of chloroplast proteins, yet the protease(s) involved is(are) not completely known. Increased expression and/or activities of serine, cysteine, aspartic, and metalloproteases were detected in senescing leaves, but these studies have not provided information on the identities of the proteases responsible for chloroplast protein breakdown. Silencing some senescence-associated proteases has delayed progression of senescence symptoms, yet it is still unclear if these proteases are directly involved in chloroplast protein breakdown. At least four cellular pathways involved in the traffic of chloroplast proteins for degradation outside the chloroplast have been described (i.e., "Rubisco-containing bodies," "senescence-associated vacuoles," "ATI1-plastid associated bodies," and "CV-containing vesicles"), which differ in their dependence on the autophagic machinery, and the identity of the proteins transported and/or degraded. Finding out the proteases involved in, for example, the degradation of Rubisco, may require piling up mutations in several senescence-associated proteases. Alternatively, targeting a proteinaceous protein inhibitor to chloroplasts may allow the inhibitor to reach "Rubisco-containing bodies," "senescence-associated vacuoles," "ATI1-plastid associated bodies," and "CV-containing vesicles" in essentially the way as chloroplast-targeted fluorescent proteins re-localize to these vesicular structures. This might help to reduce proteolytic activity, thereby reducing or slowing down plastid protein degradation during senescence.
Keywords: SAG12; chloroplast protein degradation; leaf senescence; protease; senescence-associated vacuoles; vacuole.
Figures
References
-
- Aro E. M., Virgin I., Andersson B. (1993). Photoinhibition of photosystem II. Inactivation, protein damage and turnover. Biochim. Biophys. Acta 1143, 113–134. - PubMed
Publication types
LinkOut - more resources
Full Text Sources