

Mechanisms of increased mitochondria-dependent necrosis in Wiskott-Aldrich syndrome platelets
- PMID: 31278208
- PMCID: PMC7109739
- DOI: 10.3324/haematol.2018.214460
Mechanisms of increased mitochondria-dependent necrosis in Wiskott-Aldrich syndrome platelets
Abstract
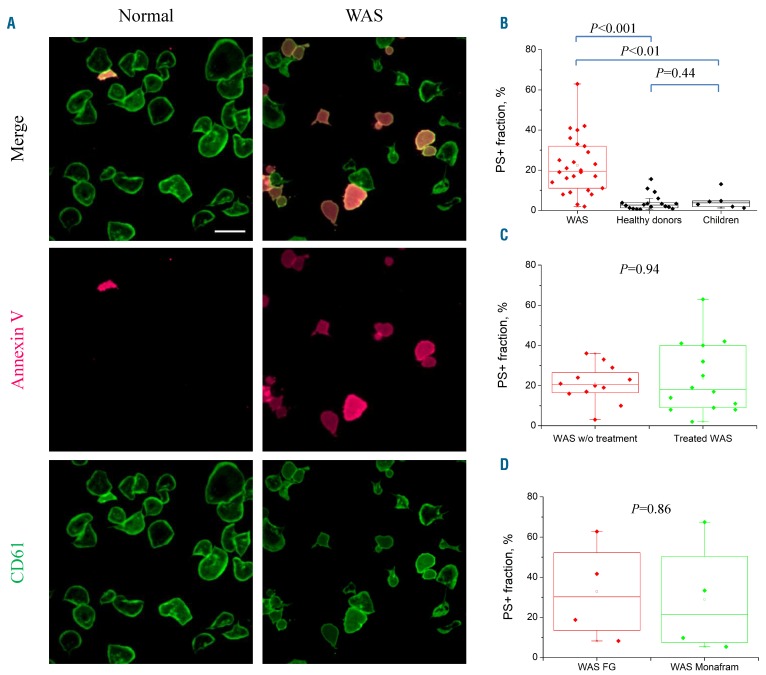
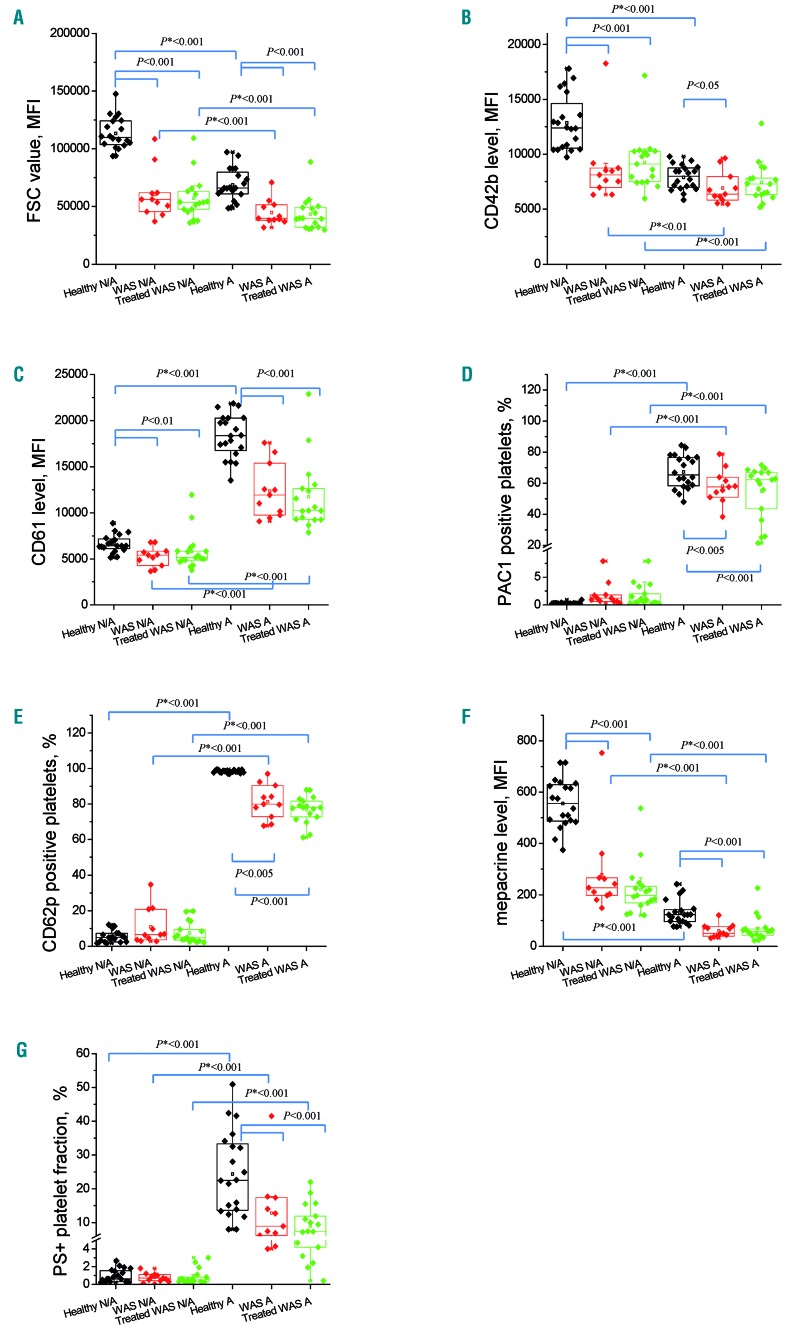
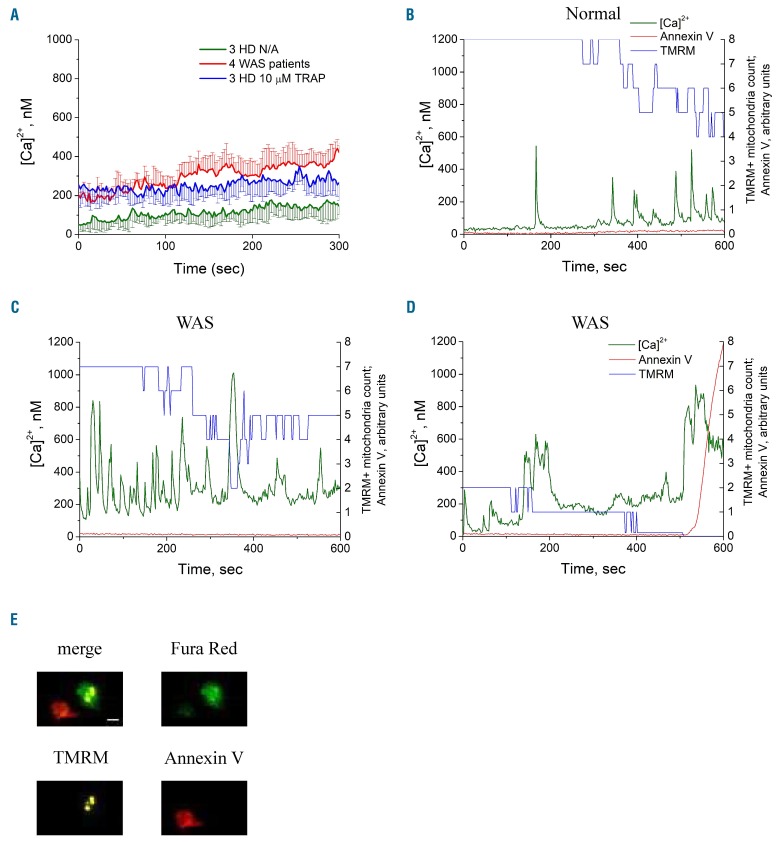
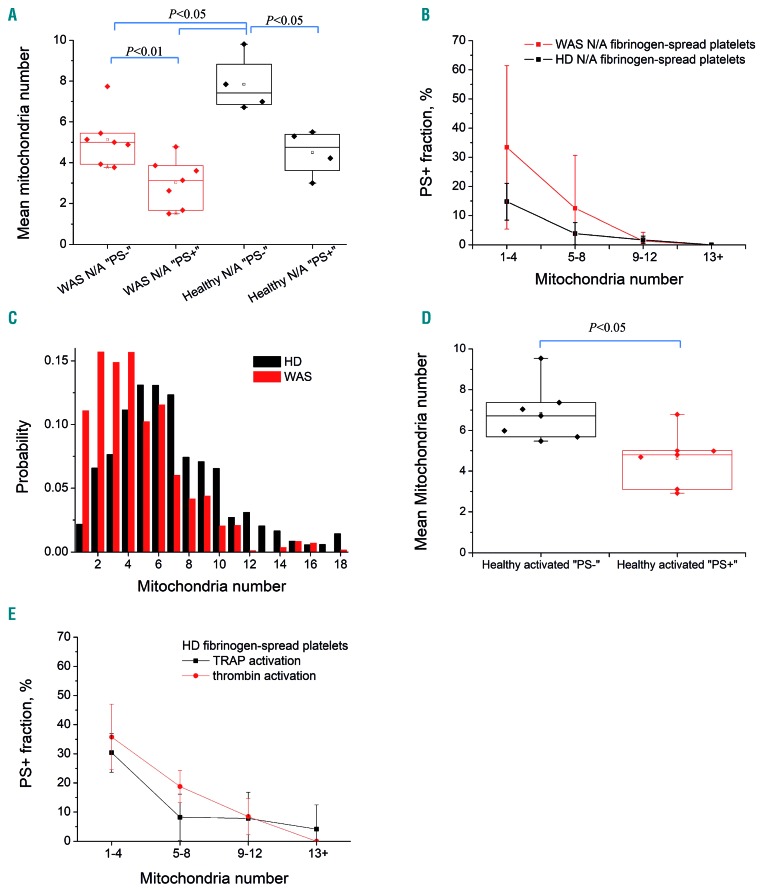
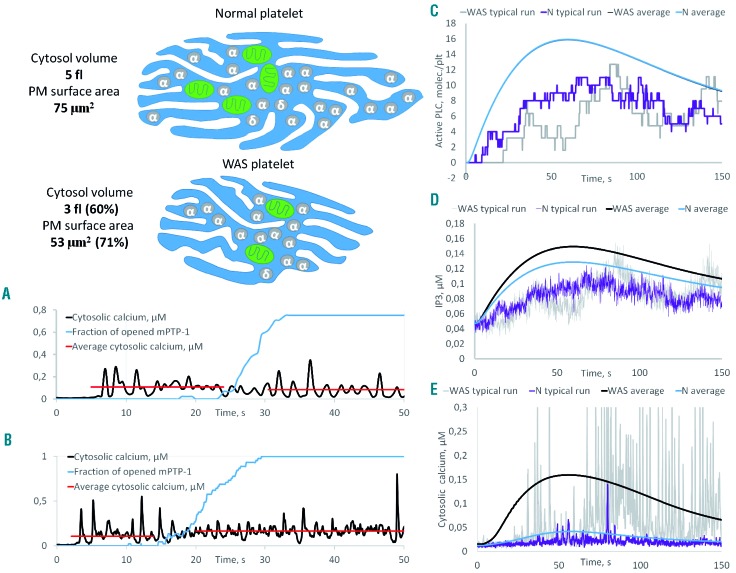
Wiskott-Aldrich syndrome (WAS) is associated with thrombocytopenia of unclear origin. We investigated real-time cytosolic calcium dynamics, mitochondrial membrane potential and phoszphatidylserine (PS) exposure in single fibrinogen-bound platelets using confocal microscopy. The WAS platelets had higher resting calcium levels, more frequent spikes, and their mitochondria more frequently lost membrane potential followed by PS exposure (in 22.9% of platelets vs 3.9% in controls; P<0.001) after the collapse of the last mitochondria. This phenomenon was inhibited by the mitochondrial permeability transition pore inhibitor cyclosporine A, as well by xestospongin C and lack of extracellular calcium. Thapsigargin by itself caused accelerated cell death in the WAS platelets. The number of mitochondria was predictive of PS exposure: 33% of platelets from WAS patients with fewer than five mitochondria exposed PS, while only 12% did among those that had five or more mitochondria. Interestingly, healthy donor platelets with fewer mitochondria also more readily became procoagulant upon PAR1/PAR4 stimulation. Collapse of single mitochondria led to greater cytosolic calcium increase in WAS platelets if they had one to three mitochondria compared with platelets containing higher numbers. A computer systems biology model of platelet calcium homeostasis showed that smaller platelets with fewer mitochondria could have impaired calcium homeostasis because of higher surface-to-volume ratio and greater metabolic load, respectively. There was a correlation (C=0.81, P<0.02) between the mean platelet size and platelet count in the WAS patients. We conclude that WAS platelets readily expose PS via a mitochondria-dependent necrotic mechanism caused by their smaller size, which could contribute to the development of thrombocytopenia.
Copyright© 2020 Ferrata Storti Foundation.
Figures

References
-
- Candotti F. Clinical manifestations and pathophysiological mechanisms of the Wiskott-Aldrich syndrome. J Clin Immunol. 2018;38(1):13–27. - PubMed
-
- Sereni L, Castiello MC, Villa A. Platelets in Wiskott-Aldrich syndrome: victims or executioners¿ J Leukoc Biol. 2018;103(3):577–590. - PubMed
-
- Kajiwara M, Nonoyama S, Eguchi M, et al. WASP is involved in proliferation and differentiation of human haemopoietic progenitors in vitro. Br J Haematol. 1999;107(2):254–262. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
