

Insulin/IGF Signaling and Vitellogenin Provisioning Mediate Intergenerational Adaptation to Nutrient Stress
- PMID: 31280992
- PMCID: PMC6650306
- DOI: 10.1016/j.cub.2019.05.062
Insulin/IGF Signaling and Vitellogenin Provisioning Mediate Intergenerational Adaptation to Nutrient Stress
Abstract
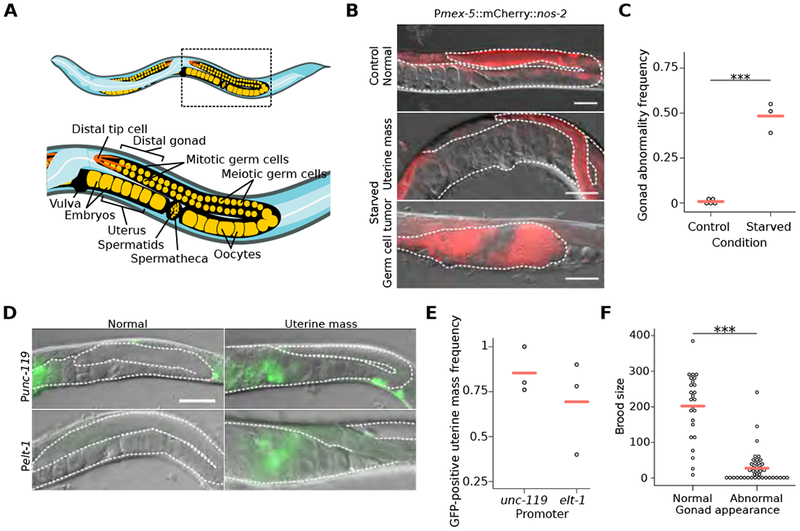
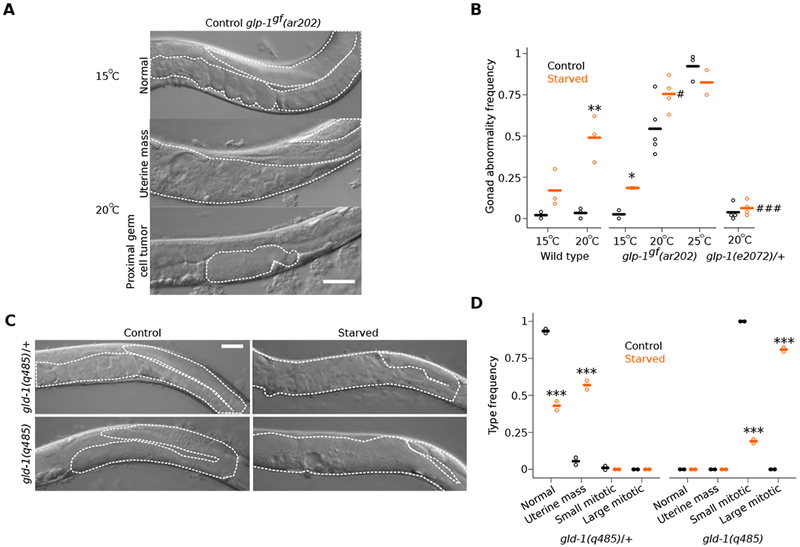
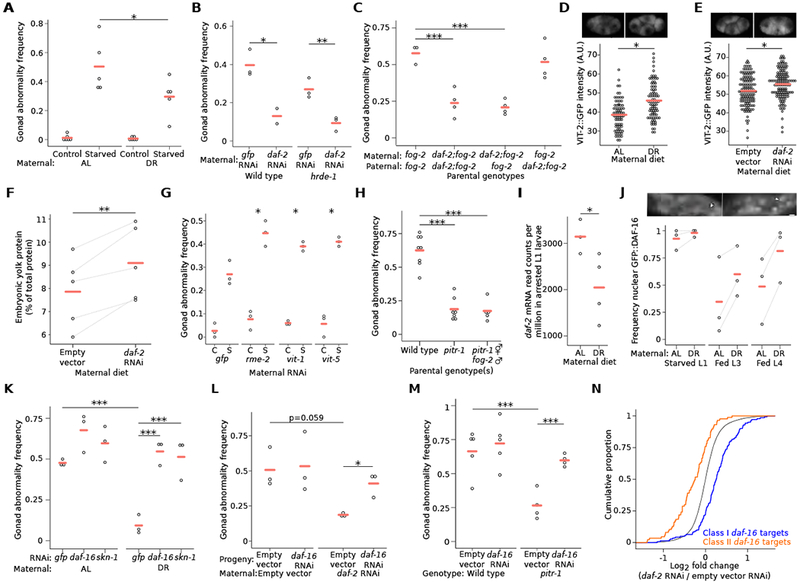
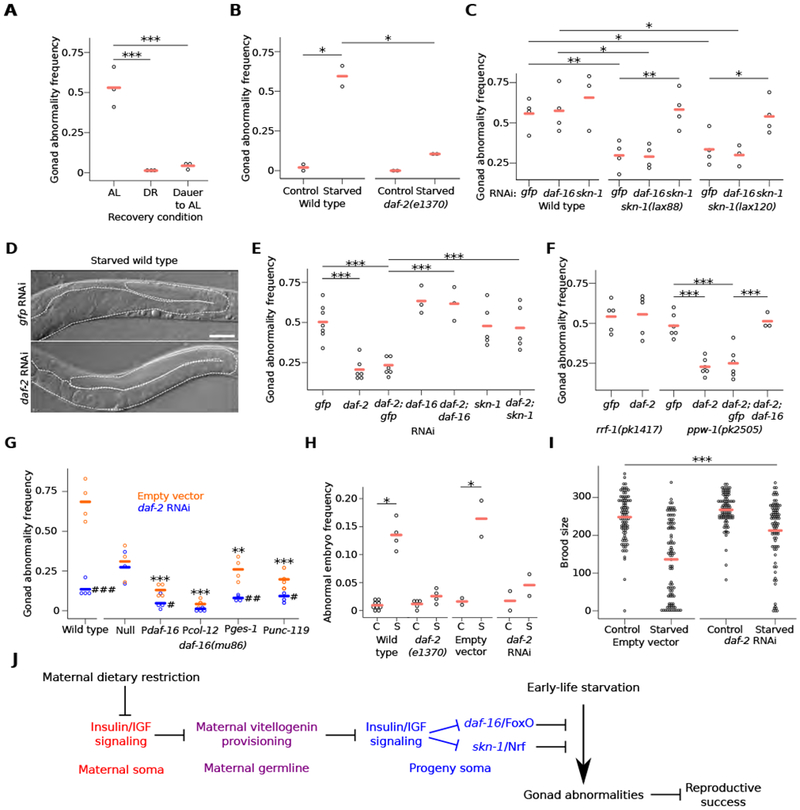
The roundworm C. elegans reversibly arrests larval development during starvation [1], but extended early-life starvation reduces reproductive success [2, 3]. Maternal dietary restriction (DR) buffers progeny from starvation as young larvae, preserving reproductive success [4]. However, the developmental basis of reduced fertility following early-life starvation is unknown, and it is unclear how maternal diet modifies developmental physiology in progeny. We show here that extended starvation in first-stage (L1) larvae followed by unrestricted feeding results in a variety of developmental abnormalities in the reproductive system, including proliferative germ-cell tumors and uterine masses that express neuronal and epidermal cell fate markers. We found that maternal DR and reduced maternal insulin/insulin-like growth factor (IGF) signaling (IIS) increase oocyte provisioning of vitellogenin lipoprotein, reducing penetrance of starvation-induced abnormalities in progeny, including tumors. Furthermore, we show that maternal DR and reduced maternal IIS reduce IIS in progeny. daf-16/FoxO and skn-1/Nrf, transcriptional effectors of IIS, are required in progeny for maternal DR and increased vitellogenin provisioning to suppress starvation-induced abnormalities. daf-16/FoxO activity in somatic tissues is sufficient to suppress starvation-induced abnormalities, suggesting cell-nonautonomous regulation of reproductive system development. This work reveals that early-life starvation compromises reproductive development and that vitellogenin-mediated intergenerational insulin/IGF-to-insulin/IGF signaling mediates adaptation to nutrient availability.
Keywords: L1 arrest; L1 diapause; dietary restriction; insulin; maternal provisioning; starvation; tumors; vitellogenin.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of Interests
Authors declare no competing interests.
Figures
Comment in
-
Maternal Inheritance: Longevity Programs Nourish Progeny via Yolk.Curr Biol. 2019 Aug 5;29(15):R748-R751. doi: 10.1016/j.cub.2019.06.050. Curr Biol. 2019. PMID: 31386852
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
