

Skp1 isoforms are differentially modified by a dual function prolyl 4-hydroxylase/N-acety lglucosaminyltransferase in a plant pathogen
- PMID: 31281925
- PMCID: PMC6774341
- DOI: 10.1093/glycob/cwz049
Skp1 isoforms are differentially modified by a dual function prolyl 4-hydroxylase/N-acety lglucosaminyltransferase in a plant pathogen
Abstract
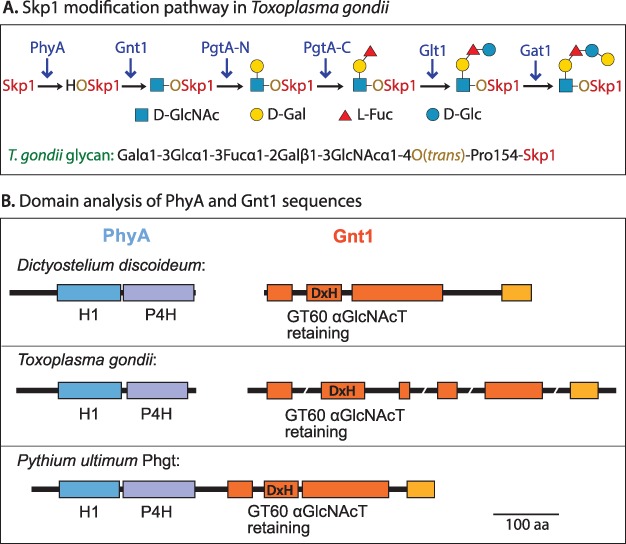
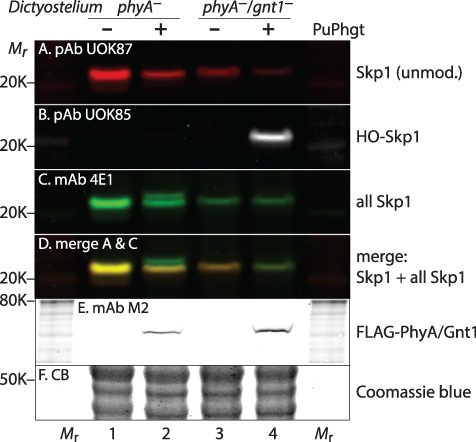
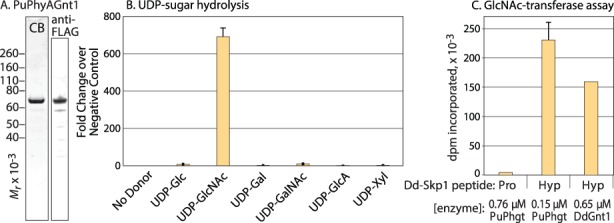
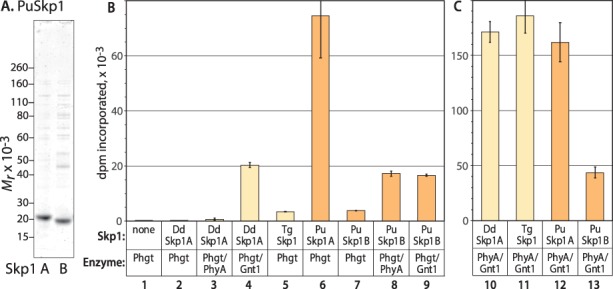
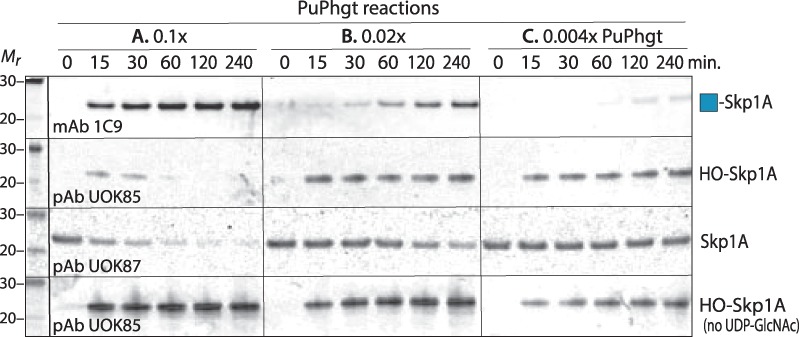
Skp1 is hydroxylated by an O2-dependent prolyl hydroxylase (PhyA) that contributes to O2-sensing in the social amoeba Dictyostelium and the mammalian pathogen Toxoplasma gondii. HO-Skp1 is subject to glycosylation and the resulting pentasaccharide affects Skp1 conformation in a way that influences association of Skp1 with F-box proteins, and potentially the assembly of E3(SCF) ubiquitin ligase complexes that mediate the polyubiquitination of target proteins that are degraded in the 26S-proteasome. To investigate the conservation and specificity of these modifications, we analyzed proteins from the oomycete Pythium ultimum, an important crop plant pathogen. Putative coding sequences for Pythium's predicted PhyA and first glycosyltransferase in the predicted five-enzyme pathway, a GlcNAc-transferase (Gnt1), predict a bifunctional enzyme (Phgt) that, when expressed in Dictyostelium, rescued a knockout of phyA but not gnt1. Though recombinant Phgt was also unable to glycosylate Dictyostelium HO-Skp1, it could hydrolyze UDP-GlcNAc and modify a synthetic hydroxypeptide from Dictyostelium Skp1. Pythium encodes two highly similar Skp1 isoforms, but only Skp1A was efficiently hydroxylated and glycosylated in vitro. While kinetic analysis revealed no evidence for processive processing of Skp1, the physical linkage of the two activities implies dedication to Skp1 in vivo. These findings indicate a widespread occurrence of the Skp1 modification pathway across protist phylogeny, suggest that both Gnt1 and PhyA are specific for Skp1 and indicate that the second Skp1 provides a bypass mechanism for O2-regulation in Pythium and other protists that conserve this gene.
Keywords: Pythium ultimum; E3 ubiquitin ligase; glycosyltransferase; plant pathogen; prolyl hydroxylase.
© The Author(s) 2019. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
References
-
- Bochkareva E, Frappier L, Edwards AM, Bochkarev A. 1998. The RPA32 subunit of human replication protein A contains a single-stranded DNA-binding domain. J Biol Chem. 273:3932–3936. - PubMed
-
- Deshaies RJ, Joazeiro CA. 2009. RING domain E3 ubiquitin ligases. Annu Rev Biochem. 78:399–434. - PubMed
-
- Heise N, Singh D, van der Wel H, Sassi SO, Johnson JM, Feasley CL, Koeller CM, Previato JO, Mendonça-Previato L, West CM. 2009. Molecular analysis of a UDP-GlcNAc:polypeptide alpha-N-acetylglucosaminyltransferase implicated in the initiation of mucin-type O-glycosylation in Trypanosoma cruzi. Glycobiology. 19:918–933. - PMC - PubMed
-
- Islam MS, Leissing TM, Chowdhury R, Hopkinson RJ, Schofield CJ. 2018. 2-oxoglutarate-dependent oxygenases. Annu Rev Biochem. 87:585–620. - PubMed
