

BODIPY-cholesterol can be reliably used to monitor cholesterol efflux from capacitating mammalian spermatozoa
- PMID: 31285440
- PMCID: PMC6614389
- DOI: 10.1038/s41598-019-45831-7
BODIPY-cholesterol can be reliably used to monitor cholesterol efflux from capacitating mammalian spermatozoa
Abstract
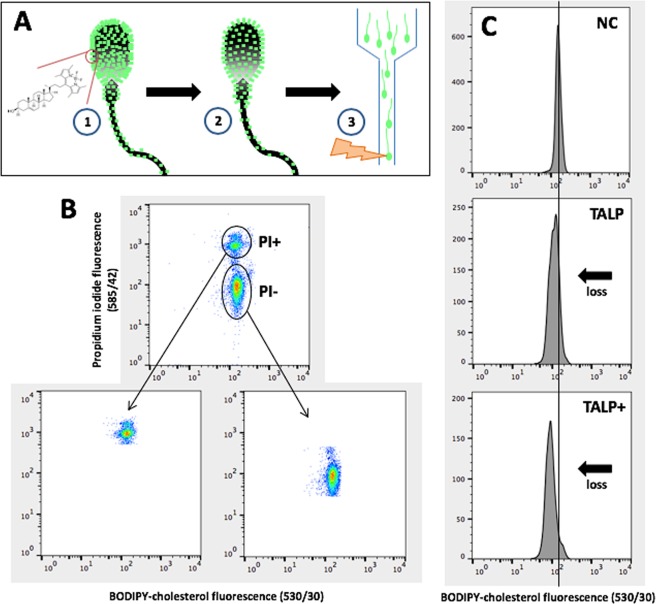
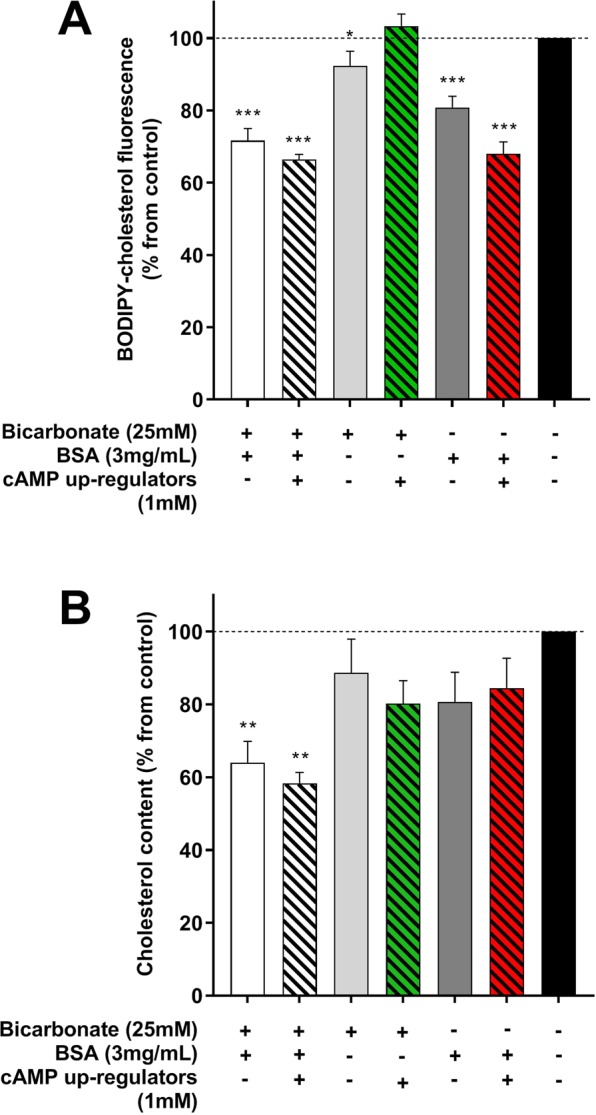
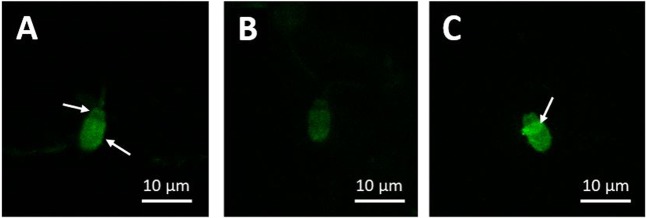
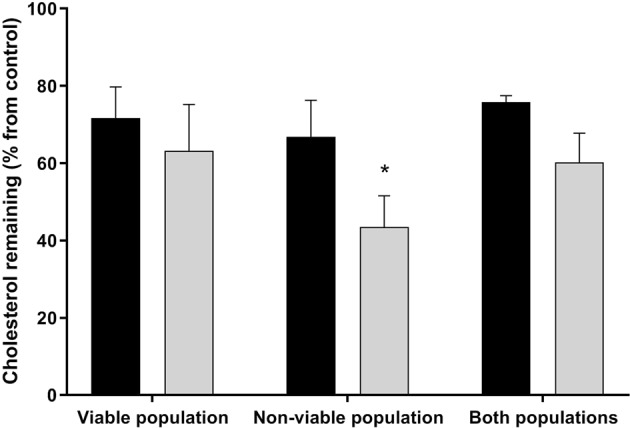
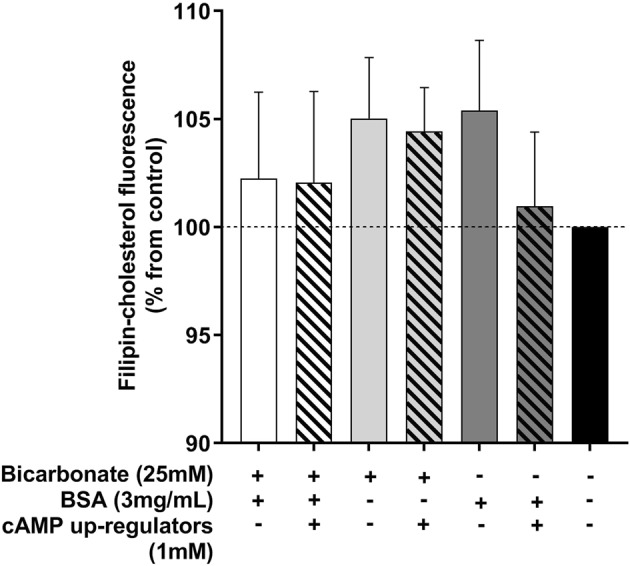
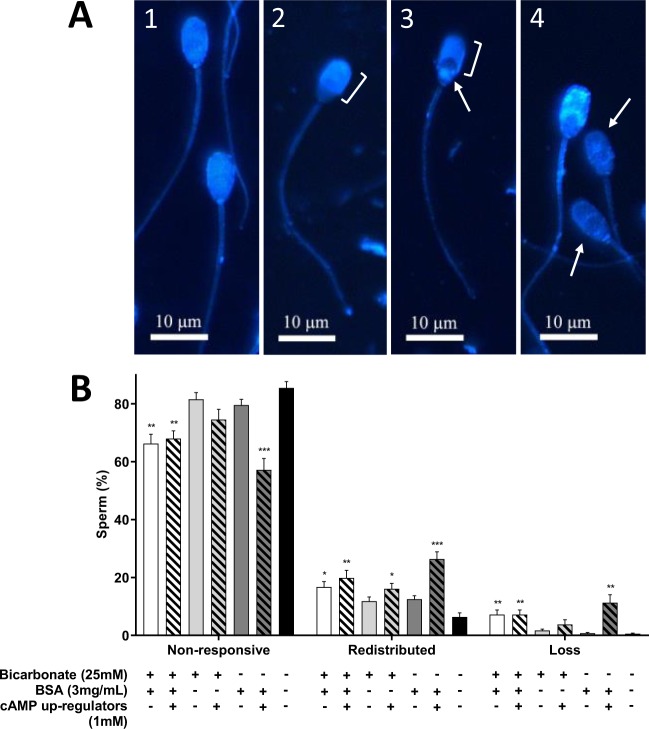
Capacitation is the final maturation step spermatozoa undergo prior to fertilisation. The efflux of cholesterol from the sperm membrane to the extracellular environment is a crucial step during capacitation but current methods to quantify this process are suboptimal. In this study, we validate the use of a BODIPY-cholesterol assay to quantify cholesterol efflux from spermatozoa during in vitro capacitation, using the boar as a model species. The novel flow cytometric BODIPY-cholesterol assay was validated with endogenous cholesterol loss as measured by mass spectrometry and compared to filipin labelling. Following exposure to a range of conditions, the BODIPY-cholesterol assay was able to detect and quantify cholesterol efflux akin to that measured with mass spectrometry. The ability to counterstain for viability is a unique feature of this assay that allowed us to highlight the importance of isolating viable cells only for a reliable measure of cholesterol efflux. Finally, the BODIPY-cholesterol assay proved to be the superior method to quantify cholesterol efflux relative to filipin labelling, though filipin remains useful for assessing cholesterol redistribution. Taken together, the BODIPY-cholesterol assay is a simple, inexpensive and reliable flow cytometric method for the measurement of cholesterol efflux from spermatozoa during in vitro capacitation.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Synergism between albumin, bicarbonate and cAMP upregulation for cholesterol efflux from ram sperm.Reproduction. 2020 Aug;160(2):269-280. doi: 10.1530/REP-19-0430. Reproduction. 2020. PMID: 32460237
-
Regionalization and redistribution of membrane phospholipids and cholesterol in mouse spermatozoa during in vitro capacitation.Biol Reprod. 1996 Nov;55(5):1133-46. doi: 10.1095/biolreprod55.5.1133. Biol Reprod. 1996. PMID: 8902227
-
Cholesterol efflux alters lipid raft stability and distribution during capacitation of boar spermatozoa.Biol Reprod. 2004 Jul;71(1):253-65. doi: 10.1095/biolreprod.103.026435. Epub 2004 Mar 17. Biol Reprod. 2004. PMID: 15028630
-
Novel methods to detect capacitation-related changes in spermatozoa.Theriogenology. 2019 Oct 1;137:56-66. doi: 10.1016/j.theriogenology.2019.05.038. Epub 2019 May 31. Theriogenology. 2019. PMID: 31230703 Review.
-
The role of cholesterol efflux in regulating the fertilization potential of mammalian spermatozoa.J Clin Invest. 2002 Sep;110(6):731-6. doi: 10.1172/JCI16392. J Clin Invest. 2002. PMID: 12235100 Free PMC article. Review. No abstract available.
Cited by
-
Time to revise: impact of methodology on boar sperm capacitation in vitro via phosphotyrosine patterns.BMC Vet Res. 2025 Jul 7;21(1):448. doi: 10.1186/s12917-025-04900-y. BMC Vet Res. 2025. PMID: 40624494 Free PMC article. Review.
-
Global Metabolomic Profiling of Host Red Blood Cells Infected with Babesia divergens Reveals Novel Antiparasitic Target Pathways.Microbiol Spectr. 2023 Feb 14;11(2):e0468822. doi: 10.1128/spectrum.04688-22. Online ahead of print. Microbiol Spectr. 2023. PMID: 36786651 Free PMC article.
-
The fate of porcine sperm CRISP2 from the perinuclear theca before and after in vitro fertilization†.Biol Reprod. 2022 Nov 14;107(5):1242-1253. doi: 10.1093/biolre/ioac169. Biol Reprod. 2022. PMID: 36054334 Free PMC article.
-
Bisphenol A Diglycidyl Ether (BADGE) and Progesterone Do Not Induce Ca2+ Signals in Boar Sperm Cells.Front Physiol. 2020 Jul 7;11:785. doi: 10.3389/fphys.2020.00785. eCollection 2020. Front Physiol. 2020. PMID: 32774306 Free PMC article.
-
Characterization of acrosin and acrosin binding protein as novel CRISP2 interacting proteins in boar spermatozoa.Andrology. 2023 Oct;11(7):1460-1471. doi: 10.1111/andr.13413. Epub 2023 Mar 9. Andrology. 2023. PMID: 36815564 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
