

The antimalarial drug amodiaquine stabilizes p53 through ribosome biogenesis stress, independently of its autophagy-inhibitory activity
- PMID: 31285544
- PMCID: PMC7205879
- DOI: 10.1038/s41418-019-0387-5
The antimalarial drug amodiaquine stabilizes p53 through ribosome biogenesis stress, independently of its autophagy-inhibitory activity
Abstract
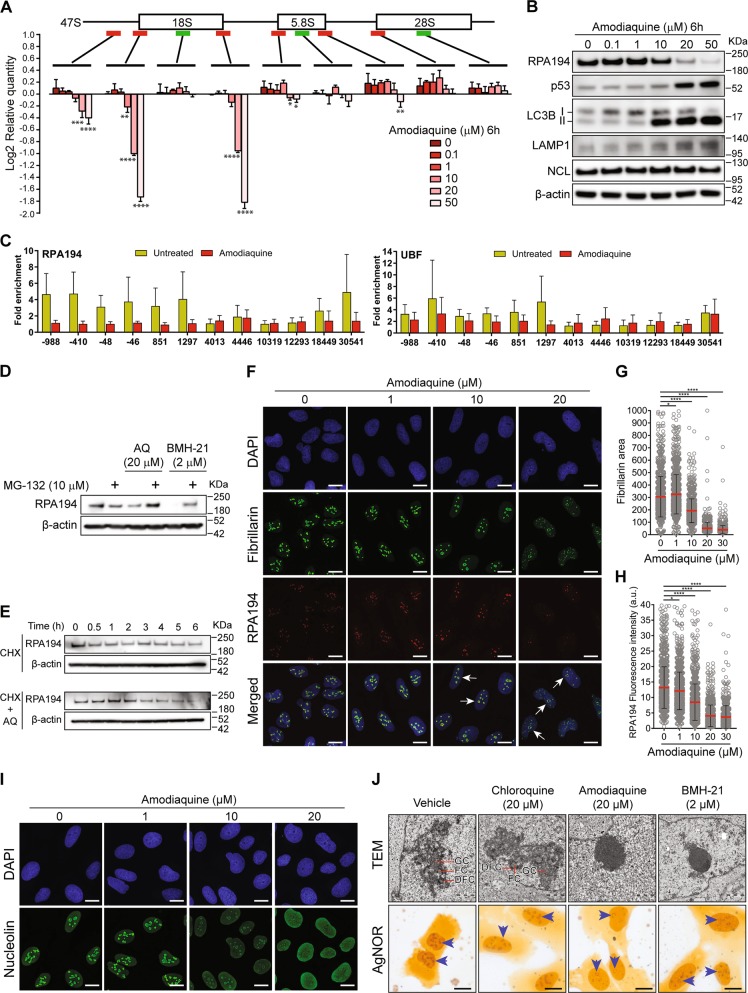
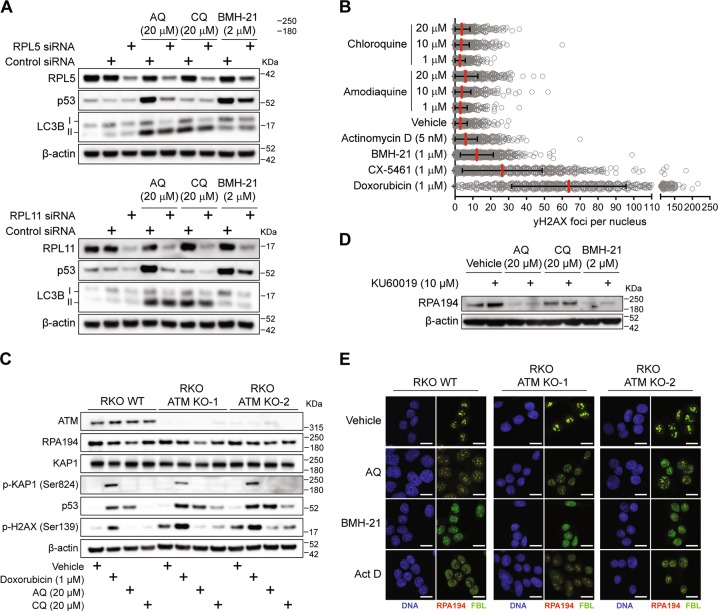
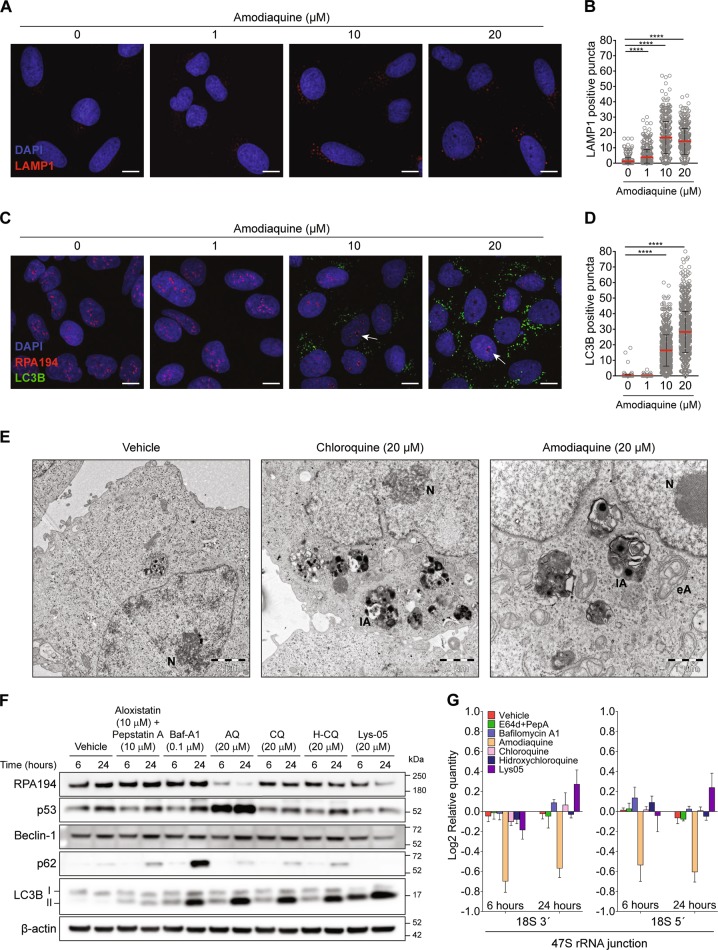
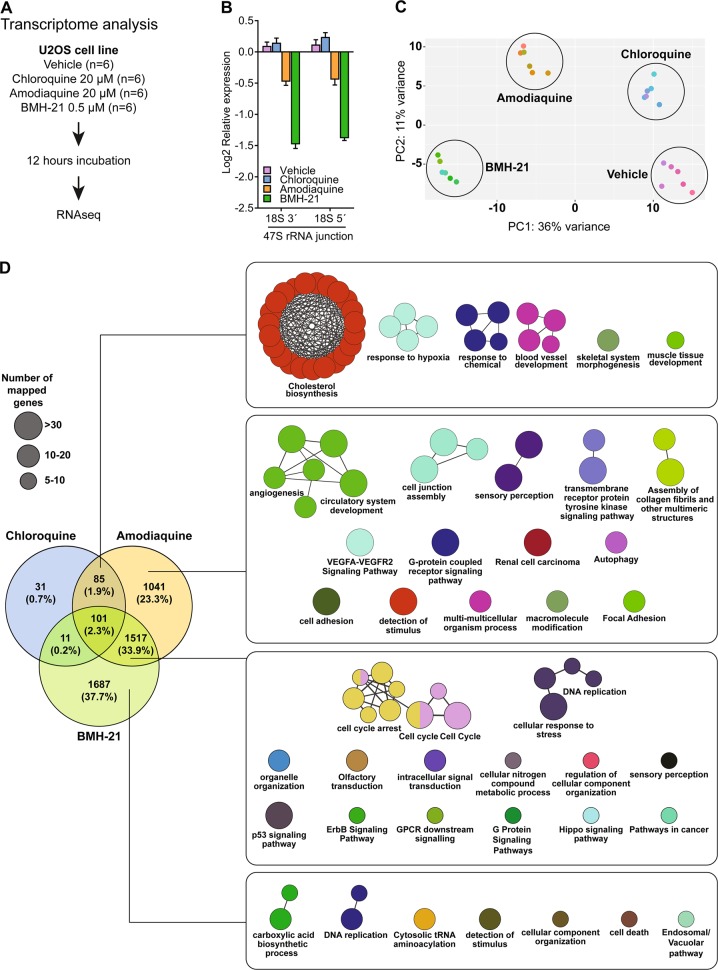
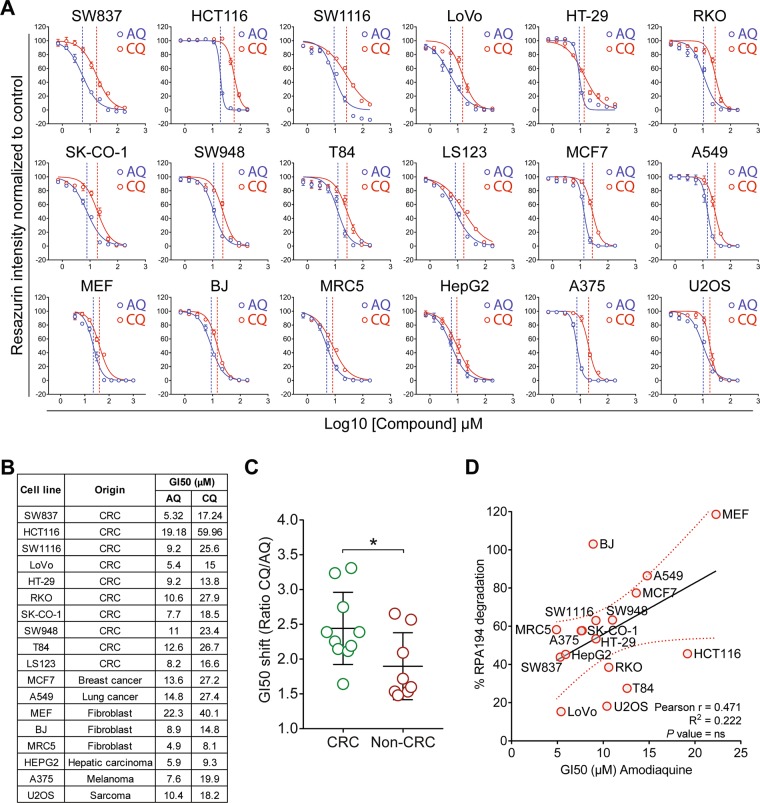
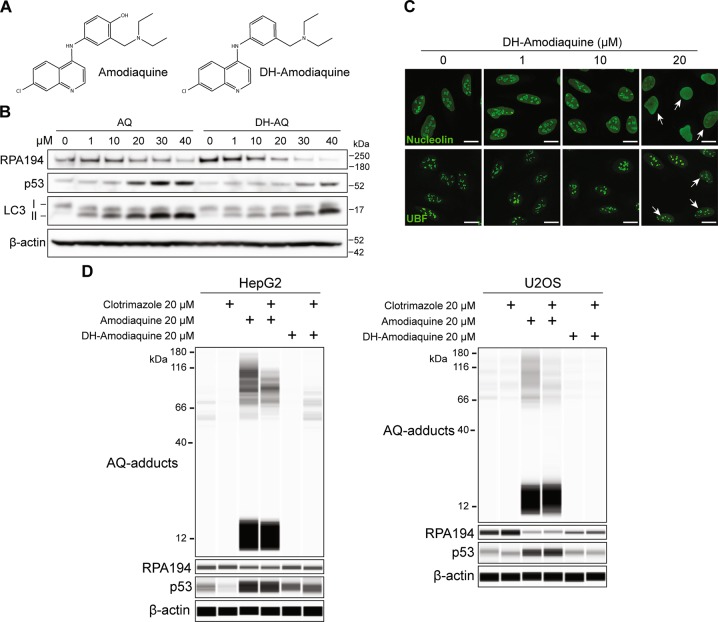
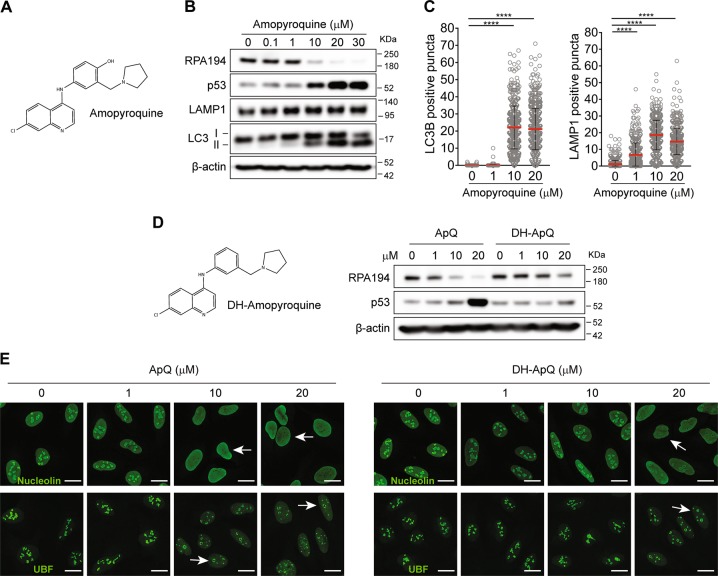
Pharmacological inhibition of ribosome biogenesis is a promising avenue for cancer therapy. Herein, we report a novel activity of the FDA-approved antimalarial drug amodiaquine which inhibits rRNA transcription, a rate-limiting step for ribosome biogenesis, in a dose-dependent manner. Amodiaquine triggers degradation of the catalytic subunit of RNA polymerase I (Pol I), with ensuing RPL5/RPL11-dependent stabilization of p53. Pol I shutdown occurs in the absence of DNA damage and without the subsequent ATM-dependent inhibition of rRNA transcription. RNAseq analysis revealed mechanistic similarities of amodiaquine with BMH-21, the first-in-class Pol I inhibitor, and with chloroquine, the antimalarial analog of amodiaquine, with well-established autophagy-inhibitory activity. Interestingly, autophagy inhibition caused by amodiaquine is not involved in the inhibition of rRNA transcription, suggesting two independent anticancer mechanisms. In vitro, amodiaquine is more efficient than chloroquine in restraining the proliferation of human cell lines derived from colorectal carcinomas, a cancer type with predicted susceptibility to ribosome biogenesis stress. Taken together, our data reveal an unsuspected activity of a drug approved and used in the clinics for over 30 years, and provide rationale for repurposing amodiaquine in cancer therapy.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
The antimalarial amodiaquine causes autophagic-lysosomal and proliferative blockade sensitizing human melanoma cells to starvation- and chemotherapy-induced cell death.Autophagy. 2013 Dec;9(12):2087-102. doi: 10.4161/auto.26506. Epub 2013 Oct 8. Autophagy. 2013. PMID: 24113242 Free PMC article.
-
Direct relationship between the level of p53 stabilization induced by rRNA synthesis-inhibiting drugs and the cell ribosome biogenesis rate.Oncogene. 2016 Feb 25;35(8):977-89. doi: 10.1038/onc.2015.147. Epub 2015 May 11. Oncogene. 2016. PMID: 25961931
-
Oncogenic MYC Induces the Impaired Ribosome Biogenesis Checkpoint and Stabilizes p53 Independent of Increased Ribosome Content.Cancer Res. 2019 Sep 1;79(17):4348-4359. doi: 10.1158/0008-5472.CAN-18-2718. Epub 2019 Jul 10. Cancer Res. 2019. PMID: 31292158
-
Activation of the tumor suppressor p53 upon impairment of ribosome biogenesis.Biochim Biophys Acta. 2014 Jun;1842(6):817-30. doi: 10.1016/j.bbadis.2013.08.014. Epub 2013 Oct 26. Biochim Biophys Acta. 2014. PMID: 24514102 Review.
-
The Ribosome Biogenesis-Cancer Connection.Cells. 2019 Jan 15;8(1):55. doi: 10.3390/cells8010055. Cells. 2019. PMID: 30650663 Free PMC article. Review.
Cited by
-
Actionable cancer vulnerability due to translational arrest, p53 aggregation and ribosome biogenesis stress evoked by the disulfiram metabolite CuET.Cell Death Differ. 2023 Jul;30(7):1666-1678. doi: 10.1038/s41418-023-01167-4. Epub 2023 May 4. Cell Death Differ. 2023. PMID: 37142656 Free PMC article.
-
The exon-junction complex helicase eIF4A3 controls cell fate via coordinated regulation of ribosome biogenesis and translational output.Sci Adv. 2021 Aug 4;7(32):eabf7561. doi: 10.1126/sciadv.abf7561. Print 2021 Aug. Sci Adv. 2021. PMID: 34348895 Free PMC article.
-
Exploring Amodiaquine's Repurposing Potential in Breast Cancer Treatment-Assessment of In-Vitro Efficacy & Mechanism of Action.Int J Mol Sci. 2022 Sep 28;23(19):11455. doi: 10.3390/ijms231911455. Int J Mol Sci. 2022. PMID: 36232751 Free PMC article.
-
Therapeutic targeting of CNBP phase separation inhibits ribosome biogenesis and neuroblastoma progression via modulating SWI/SNF complex activity.Clin Transl Med. 2023 Apr;13(4):e1235. doi: 10.1002/ctm2.1235. Clin Transl Med. 2023. PMID: 37186134 Free PMC article.
-
Block of Proliferation 1 Promotes Proliferation, Invasion and Epithelial Mesenchymal Transformation in Gastric Cancer.Oxid Med Cell Longev. 2022 Feb 16;2022:2946989. doi: 10.1155/2022/2946989. eCollection 2022. Oxid Med Cell Longev. 2022. PMID: 35222794 Free PMC article.
References
-
- Pelletier J, Thomas G, Volarevic S. Ribosome biogenesis in cancer: new players and therapeutic avenues. Nat Rev Cancer. 2018;18:51–63. - PubMed
-
- Derenzini M, Montanaro L, Trere D. What the nucleolus says to a tumour pathologist. Histopathology. 2009;54:753–62. - PubMed
-
- Zhao J, Yuan X, Frodin M, Grummt I. ERK-dependent phosphorylation of the transcription initiation factor TIF-IA is required for RNA polymerase I transcription and cell growth. Mol Cell. 2003;11:405–13. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
