

Matrix stiffness induces a tumorigenic phenotype in mammary epithelium through changes in chromatin accessibility
- PMID: 31285581
- PMCID: PMC6899165
- DOI: 10.1038/s41551-019-0420-5
Matrix stiffness induces a tumorigenic phenotype in mammary epithelium through changes in chromatin accessibility
Abstract
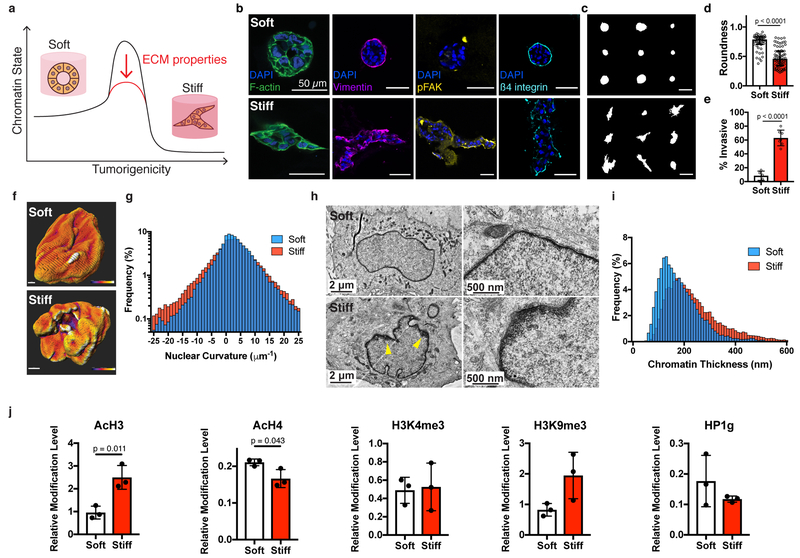
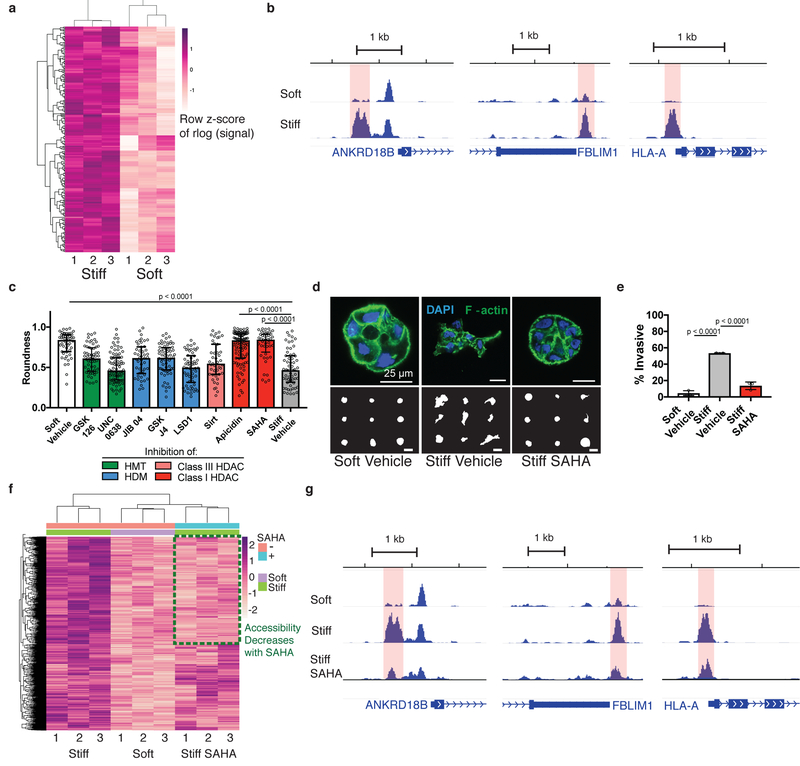
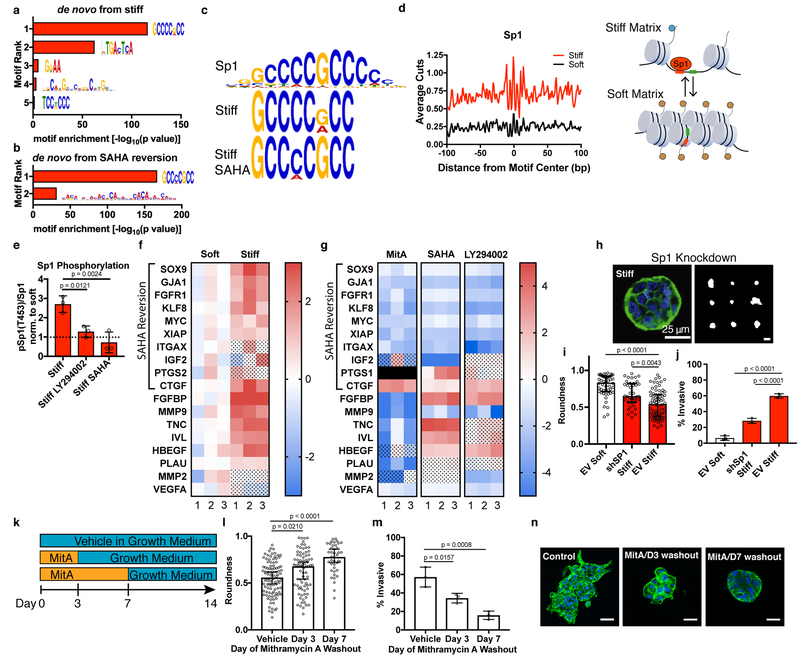
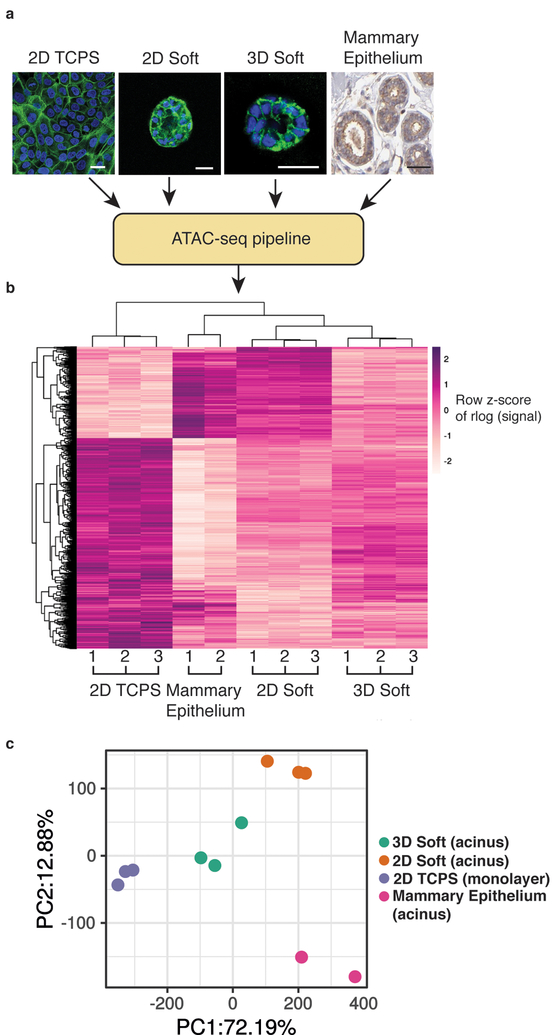
In breast cancer, the increased stiffness of the extracellular matrix is a key driver of malignancy. Yet little is known about the epigenomic changes that underlie the tumorigenic impact of extracellular matrix mechanics. Here, we show in a three-dimensional culture model of breast cancer that stiff extracellular matrix induces a tumorigenic phenotype through changes in chromatin state. We found that increased stiffness yielded cells with more wrinkled nuclei and with increased lamina-associated chromatin, that cells cultured in stiff matrices displayed more accessible chromatin sites, which exhibited footprints of Sp1 binding, and that this transcription factor acts along with the histone deacetylases 3 and 8 to regulate the induction of stiffness-mediated tumorigenicity. Just as cell culture on soft environments or in them rather than on tissue-culture plastic better recapitulates the acinar morphology observed in mammary epithelium in vivo, mammary epithelial cells cultured on soft microenvironments or in them also more closely replicate the in vivo chromatin state. Our results emphasize the importance of culture conditions for epigenomic studies, and reveal that chromatin state is a critical mediator of mechanotransduction.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures

Comment in
-
Facing up to a hard truth.Nat Rev Cancer. 2019 Oct;19(10):542-543. doi: 10.1038/s41568-019-0195-8. Nat Rev Cancer. 2019. PMID: 31409914 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
