

24(S)-Hydroxycholesterol induces ER dysfunction-mediated unconventional cell death
- PMID: 31285856
- PMCID: PMC6611791
- DOI: 10.1038/s41420-019-0192-4
24(S)-Hydroxycholesterol induces ER dysfunction-mediated unconventional cell death
Abstract
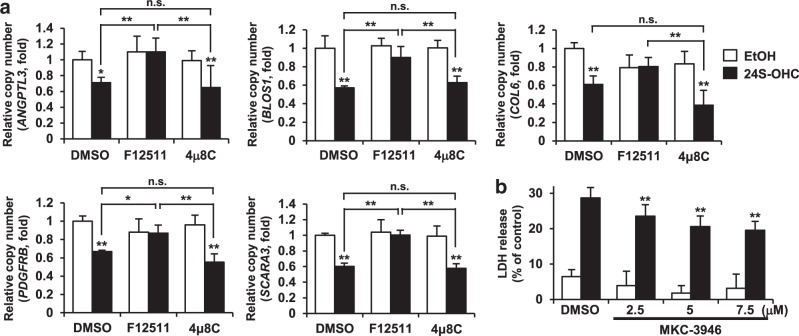
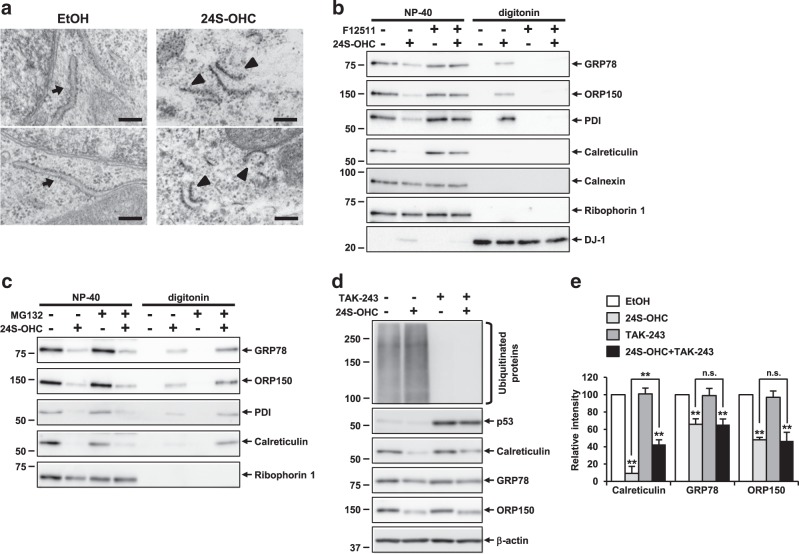
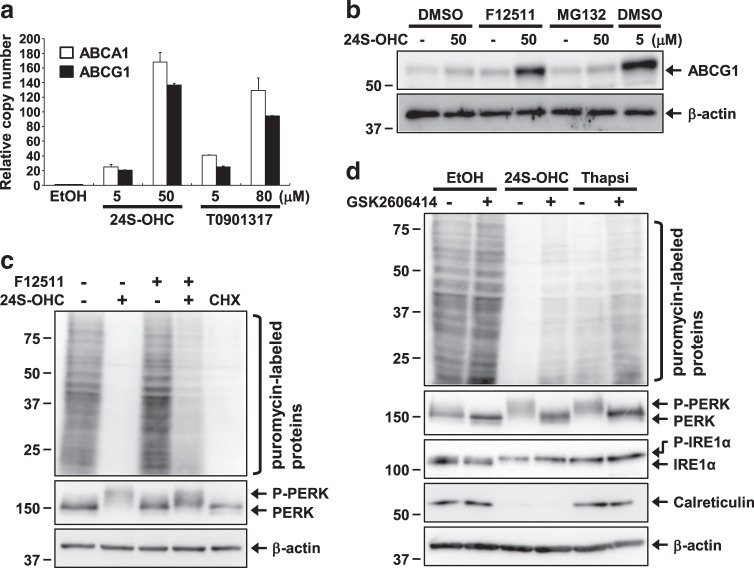
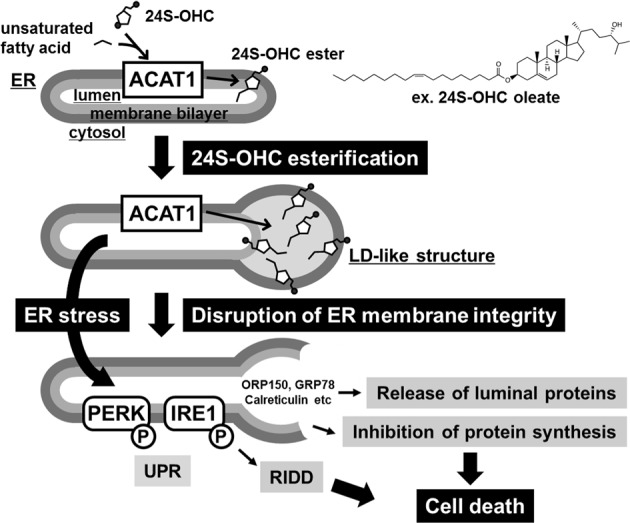
Endoplasmic reticulum (ER) stress induced by disruption of protein folding activates the unfolded protein response (UPR), which while generally pro-survival in effect can also induce cell death under severe ER stress. 24(S)-hydroxycholesterol (24S-OHC), which is enzymatically produced in the ER of neurons, plays an important role in maintaining brain cholesterol homeostasis but also shows neurotoxicity when subjected to esterification by acyl-CoA:cholesterol acyltransferase 1 (ACAT1) in the ER. In this study, we demonstrated that the accumulation of 24S-OHC esters in human neuroblastoma SH-SY5Y cells evoked the UPR with substantially no pro-survival adaptive response but with significant activation of pro-death UPR signaling via regulated IRE1-dependent decay (RIDD). We further found that accumulation of 24S-OHC esters caused disruption of ER membrane integrity and release of ER luminal proteins into cytosol. We also found that de novo synthesis of global proteins was robustly suppressed in 24S-OHC-treated cells. Collectively, these results show that ER dysfunction and the accompanying RIDD-mediated pro-death UPR signaling and global protein synthesis inhibition are responsible for 24S-OHC ester-induced unconventional cell death.
Keywords: Apoptosis; Sterols.
Conflict of interest statement
Conflict of interestThe authors declare that they have no conflict of interest.
Figures


Similar articles
-
Integrated stress response is involved in the 24(S)-hydroxycholesterol-induced unconventional cell death mechanism.Cell Death Discov. 2022 Oct 4;8(1):406. doi: 10.1038/s41420-022-01197-w. Cell Death Discov. 2022. PMID: 36195595 Free PMC article.
-
Induction of apoptosis and necroptosis by 24(S)-hydroxycholesterol is dependent on activity of acyl-CoA:cholesterol acyltransferase 1.Cell Death Dis. 2014 Jan 9;5(1):e990. doi: 10.1038/cddis.2013.524. Cell Death Dis. 2014. PMID: 24407243 Free PMC article.
-
Esterification of 24S-OHC induces formation of atypical lipid droplet-like structures, leading to neuronal cell death.J Lipid Res. 2016 Nov;57(11):2005-2014. doi: 10.1194/jlr.M068775. Epub 2016 Sep 19. J Lipid Res. 2016. PMID: 27647838 Free PMC article.
-
Different functions of vitamin E homologues in the various types of cell death induced by oxysterols.Free Radic Biol Med. 2021 Nov 20;176:356-365. doi: 10.1016/j.freeradbiomed.2021.10.008. Epub 2021 Oct 12. Free Radic Biol Med. 2021. PMID: 34648906 Review.
-
Diverse functions of 24(S)-hydroxycholesterol in the brain.Biochem Biophys Res Commun. 2014 Apr 11;446(3):692-6. doi: 10.1016/j.bbrc.2014.02.010. Epub 2014 Feb 12. Biochem Biophys Res Commun. 2014. PMID: 24530911 Review.
Cited by
-
Impact of Oxysterols on Cell Death, Proliferation, and Differentiation Induction: Current Status.Cells. 2021 Sep 3;10(9):2301. doi: 10.3390/cells10092301. Cells. 2021. PMID: 34571949 Free PMC article. Review.
-
Curcumin Derivative GT863 Inhibits Amyloid-Beta Production via Inhibition of Protein N-Glycosylation.Cells. 2020 Feb 3;9(2):349. doi: 10.3390/cells9020349. Cells. 2020. PMID: 32028683 Free PMC article.
-
The Role of Cholesterol in α-Synuclein and Lewy Body Pathology in GBA1 Parkinson's Disease.Mov Disord. 2021 May;36(5):1070-1085. doi: 10.1002/mds.28396. Epub 2020 Nov 21. Mov Disord. 2021. PMID: 33219714 Free PMC article. Review.
-
Suppression of Amyloid-β Adsorption on Endoplasmic Reticulum Stress-Mimicking Membranes by α-Tocopherol and α-Tocotrienol.J Phys Chem Lett. 2022 Dec 29;13(51):11955-11960. doi: 10.1021/acs.jpclett.2c03098. Epub 2022 Dec 19. J Phys Chem Lett. 2022. PMID: 36534445 Free PMC article.
-
Integrated stress response is involved in the 24(S)-hydroxycholesterol-induced unconventional cell death mechanism.Cell Death Discov. 2022 Oct 4;8(1):406. doi: 10.1038/s41420-022-01197-w. Cell Death Discov. 2022. PMID: 36195595 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Miscellaneous
