

Spermine in semen of male sea lamprey acts as a sex pheromone
- PMID: 31287811
- PMCID: PMC6615597
- DOI: 10.1371/journal.pbio.3000332
Spermine in semen of male sea lamprey acts as a sex pheromone
Abstract
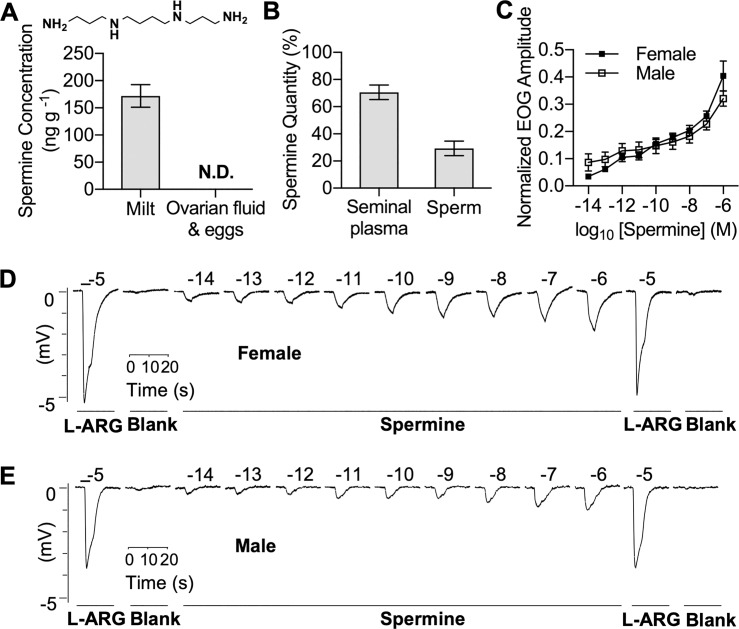
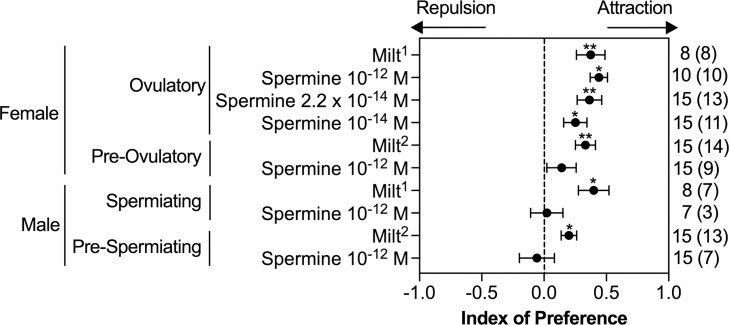
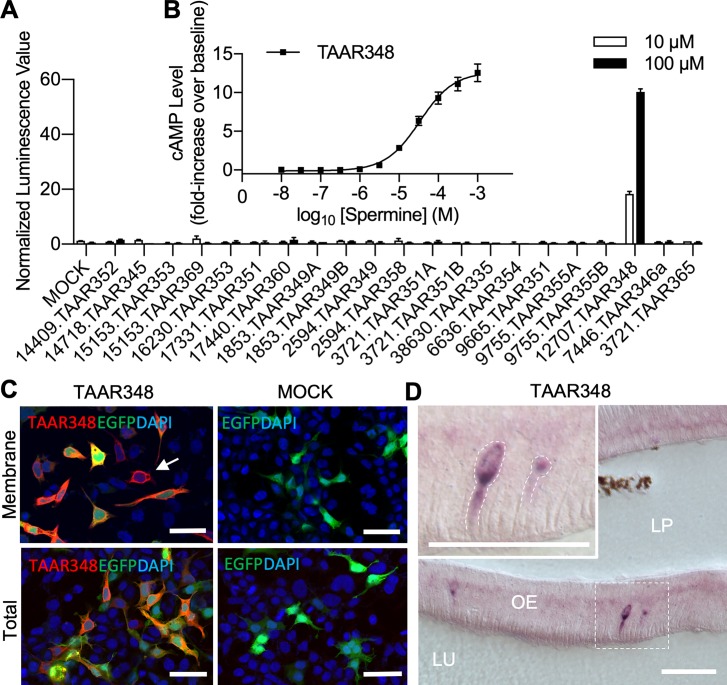
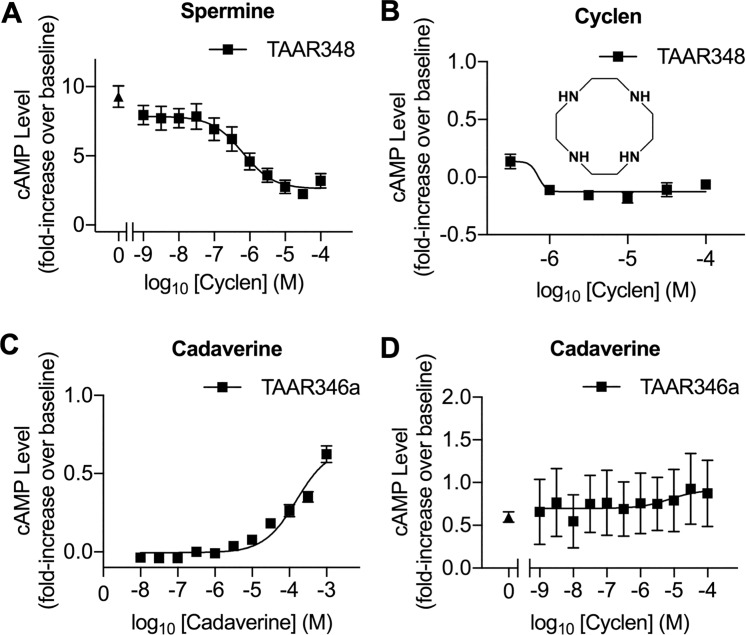
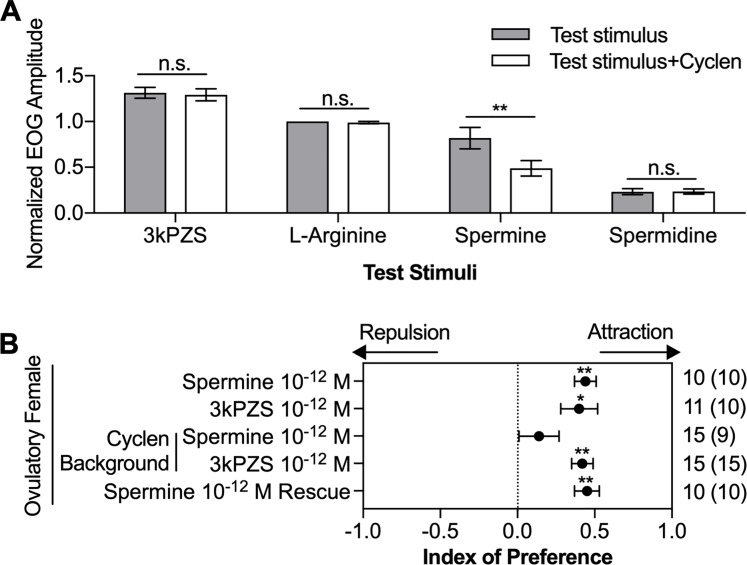
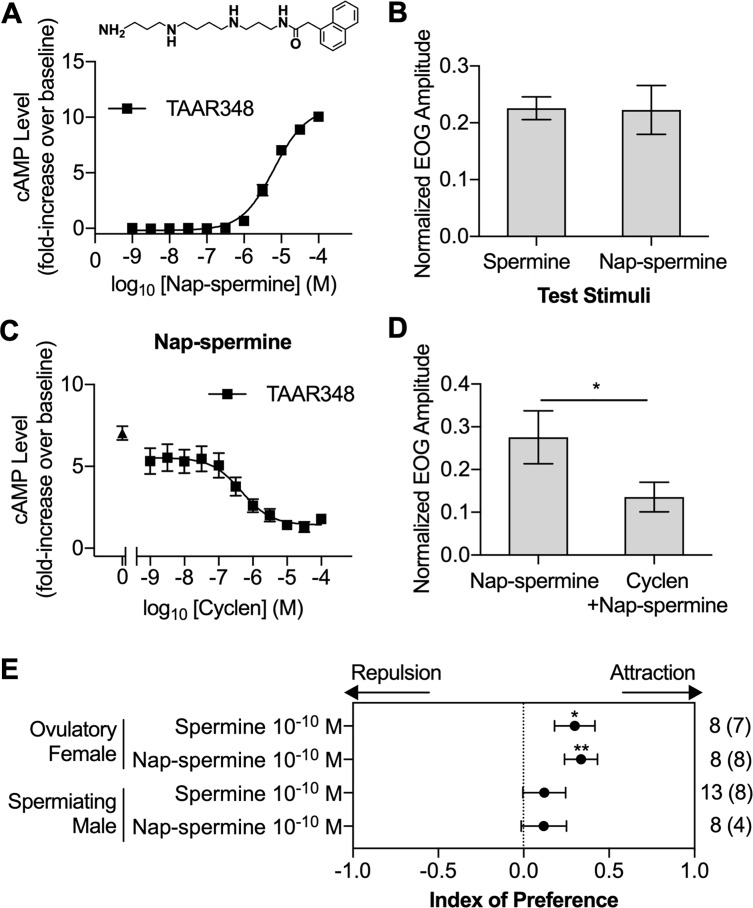
Semen is fundamental for sexual reproduction. The non-sperm part of ejaculated semen, or seminal plasma, facilitates the delivery of sperm to the eggs. The seminal plasma of some species with internal fertilization contains anti-aphrodisiac molecules that deter promiscuity in post-copulatory females, conferring fitness benefits to the ejaculating male. By contrast, in some taxa with external fertilization such as fish, exposure to semen promotes spawning behaviors. However, no specific compounds in semen have been identified as aphrodisiac pheromones. We sought to identify a pheromone from the milt (fish semen) of sea lamprey (Petromyzon marinus), a jawless fish that spawns in lek-like aggregations in which each spermiating male defends a nest, and ovulatory females move from nest to nest to mate. We postulated that milt compounds signal to ovulatory females the presence of spawning spermiating males. We determined that spermine, an odorous polyamine initially identified from human semen, is indeed a milt pheromone. At concentrations as low as 10-14 molar, spermine stimulated the lamprey olfactory system and attracted ovulatory females but did not attract males or pre-ovulatory females. We found spermine activated a trace amine-associated receptor (TAAR)-like receptor in the lamprey olfactory epithelium. A novel antagonist to that receptor nullified the attraction of ovulatory females to spermine. Our results elucidate a mechanism whereby a seminal plasma pheromone attracts ready-to-mate females and implicates a possible conservation of the olfactory detection of semen from jawless vertebrates to humans. Milt pheromones may also have management implications for sea lamprey populations.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
A Sea Lamprey (Petromyzon marinus) Sex Pheromone Mixture Increases Trap Catch Relative to a Single Synthesized Component in Specific Environments.J Chem Ecol. 2015 Mar;41(3):311-21. doi: 10.1007/s10886-015-0561-2. Epub 2015 Mar 22. J Chem Ecol. 2015. PMID: 25795091
-
Evidence that progestins play an important role in spermiation and pheromone production in male sea lamprey (Petromyzon marinus).Gen Comp Endocrinol. 2015 Feb 1;212:17-27. doi: 10.1016/j.ygcen.2015.01.008. Epub 2015 Jan 23. Gen Comp Endocrinol. 2015. PMID: 25623147
-
Effects of sex pheromones and sexual maturation on locomotor activity in female sea lamprey (Petromyzon marinus).J Biol Rhythms. 2013 Jun;28(3):218-26. doi: 10.1177/0748730413488994. J Biol Rhythms. 2013. PMID: 23735501
-
Chemical cues and pheromones in the sea lamprey (Petromyzon marinus).Front Zool. 2015 Nov 25;12:32. doi: 10.1186/s12983-015-0126-9. eCollection 2015. Front Zool. 2015. PMID: 26609313 Free PMC article. Review.
-
Petromyzonol sulfate and its derivatives: the chemoattractants of the sea lamprey.Bioessays. 2005 Feb;27(2):222-8. doi: 10.1002/bies.20155. Bioessays. 2005. PMID: 15666352 Review.
Cited by
-
Odorant Receptor OR2C1 Is an Essential Modulator of Boar Sperm Capacitation by Binding with Heparin.Int J Mol Sci. 2023 Jan 14;24(2):1664. doi: 10.3390/ijms24021664. Int J Mol Sci. 2023. PMID: 36675176 Free PMC article.
-
Non-targeted metabolomics aids in sex pheromone identification: a proof-of-concept study with the triangulate cobweb spider, Steatoda triangulosa.Sci Rep. 2023 Oct 27;13(1):18426. doi: 10.1038/s41598-023-44948-0. Sci Rep. 2023. PMID: 37891331 Free PMC article.
-
Impact of starvation on paternal reproductive investment in Neoseiulus californicus.Exp Appl Acarol. 2022 May;87(1):1-18. doi: 10.1007/s10493-022-00717-8. Epub 2022 Jun 2. Exp Appl Acarol. 2022. PMID: 35653071
-
Evolution of Brain-Expressed Biogenic Amine Receptors into Olfactory Trace Amine-Associated Receptors.Mol Biol Evol. 2022 Mar 2;39(3):msac006. doi: 10.1093/molbev/msac006. Mol Biol Evol. 2022. PMID: 35021231 Free PMC article.
-
Synergistic behavioral antagonists of a sex pheromone reduce reproduction of invasive sea lamprey.iScience. 2023 Aug 26;26(10):107744. doi: 10.1016/j.isci.2023.107744. eCollection 2023 Oct 20. iScience. 2023. PMID: 37810212 Free PMC article.
References
-
- Suarez S, Wolfner M. Seminal Plasma Plays Important Roles in Fertility In: Jonge CD, Barratt C, editors. The Sperm Cell: Production, Maturation, Fertilization, Regeneration. Cambridge: Cambridge University Press; 2017. p. 88–108.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
