

Reconstitution of Microtubule Nucleation In Vitro Reveals Novel Roles for Mzt1
- PMID: 31287970
- PMCID: PMC6616311
- DOI: 10.1016/j.cub.2019.05.058
Reconstitution of Microtubule Nucleation In Vitro Reveals Novel Roles for Mzt1
Erratum in
-
Reconstitution of Microtubule Nucleation In Vitro Reveals Novel Roles for Mzt1.Curr Biol. 2023 May 8;33(9):1864. doi: 10.1016/j.cub.2023.04.030. Curr Biol. 2023. PMID: 37160082 Free PMC article. No abstract available.
Abstract
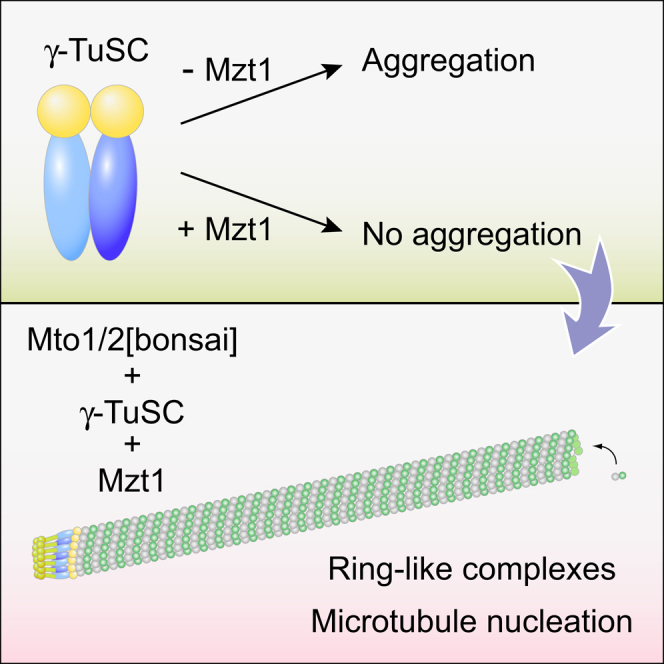
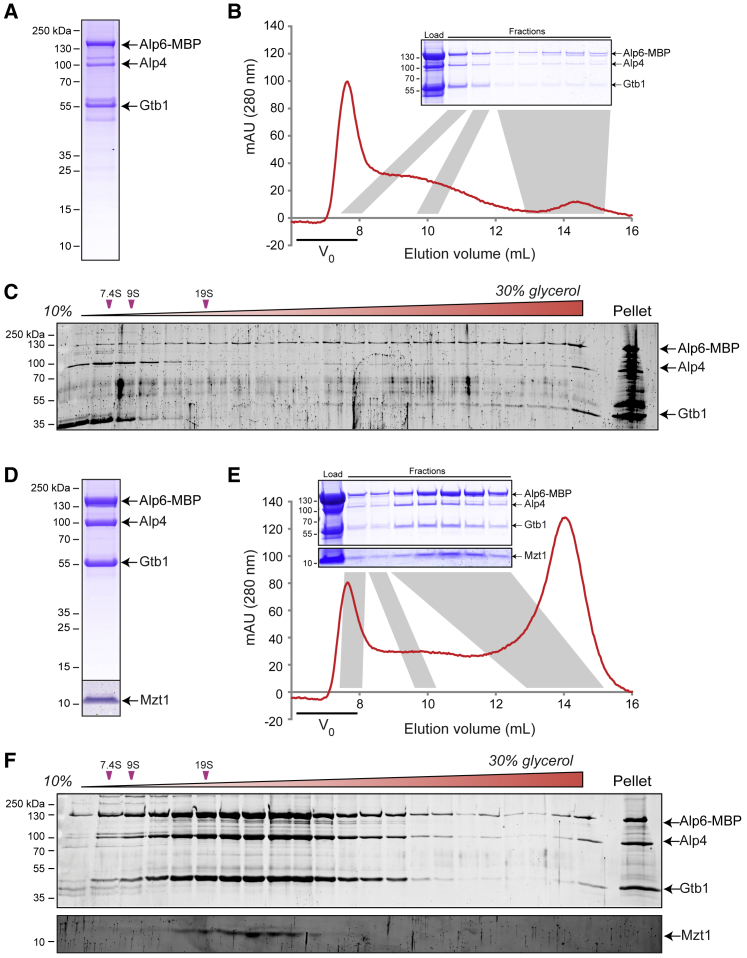
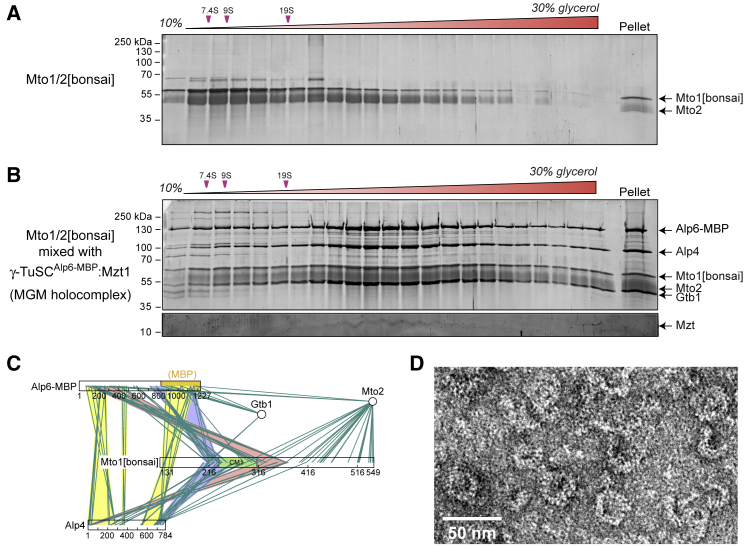
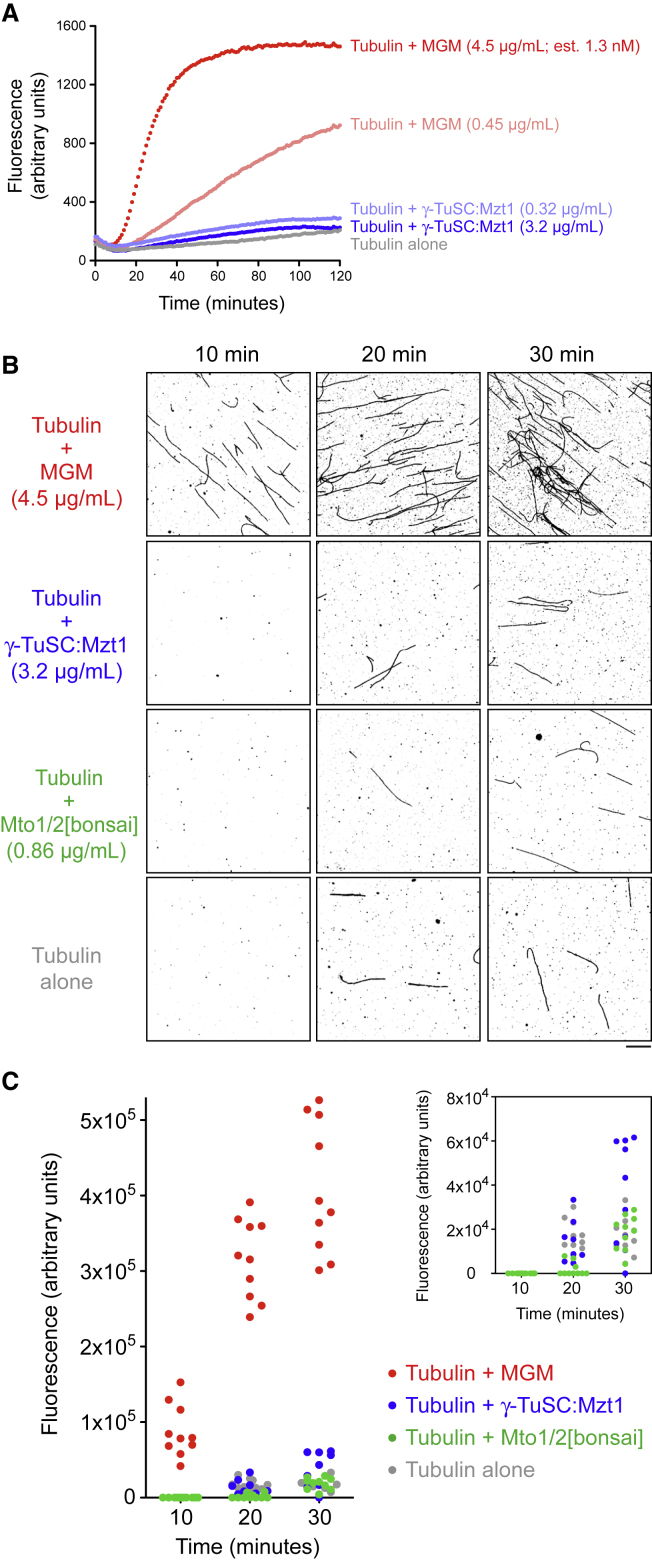
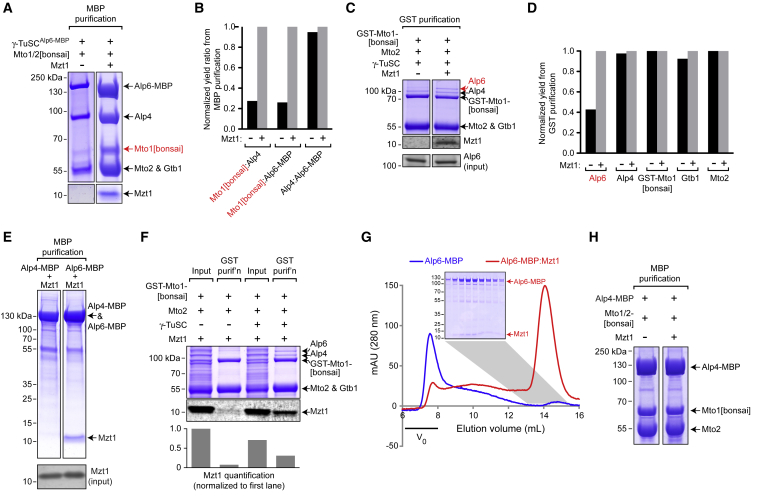
Microtubule (MT) nucleation depends on the γ-tubulin complex (γ-TuC), in which multiple copies of the heterotetrameric γ-tubulin small complex (γ-TuSC) associate to form a ring-like structure (in metazoans, γ-tubulin ring complex; γ-TuRC) [1-7]. Additional conserved regulators of the γ-TuC include the small protein Mzt1 (MOZART1 in human; GIP1/1B and GIP2/1A in plants) [8-13] and proteins containing a Centrosomin Motif 1 (CM1) domain [10, 14-19]. Many insights into γ-TuC regulators have come from in vivo analysis in fission yeast Schizosaccharomyces pombe. The S. pombe CM1 protein Mto1 recruits the γ-TuC to microtubule-organizing centers (MTOCs) [14, 20-22], and analysis of Mto1[bonsai], a truncated version of Mto1 that cannot localize to MTOCs, has shown that Mto1 also has a role in γ-TuC activation [23]. S. pombe Mzt1 interacts with γ-TuSC and is essential for γ-TuC function and localization to MTOCs [11, 12]. However, the mechanisms by which Mzt1 functions remain unclear. Here we describe reconstitution of MT nucleation using purified recombinant Mto1[bonsai], the Mto1 partner protein Mto2, γ-TuSC, and Mzt1. Multiple copies of the six proteins involved coassemble to form a 34-40S ring-like "MGM" holocomplex that is a potent MT nucleator in vitro. Using purified MGM and subcomplexes, we investigate the role of Mzt1 in MT nucleation. Our results suggest that Mzt1 is critical to stabilize Alp6, the S. pombe homolog of human γ-TuSC protein GCP3, in an "interaction-competent" form within the γ-TuSC. This is essential for MGM to become a functional nucleator.
Copyright © 2019 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Kollman J.M., Merdes A., Mourey L., Agard D.A. Microtubule nucleation by γ-tubulin complexes. Nat. Rev. Mol. Cell Biol. 2011;12:709–721. - PMC - PubMed
- Kollman, J.M., Merdes, A., Mourey, L., and Agard, D.A. (2011). Microtubule nucleation by γ-tubulin complexes. Nat. Rev. Mol. Cell Biol. 12, 709-721. - PMC - PubMed
-
- Lin T.C., Neuner A., Schiebel E. Targeting of γ-tubulin complexes to microtubule organizing centers: conservation and divergence. Trends Cell Biol. 2015;25:296–307. - PubMed
- Lin, T.C., Neuner, A., and Schiebel, E. (2015). Targeting of γ-tubulin complexes to microtubule organizing centers: conservation and divergence. Trends Cell Biol. 25, 296-307. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
