

Bumblebees learn foraging routes through exploitation-exploration cycles
- PMID: 31288648
- PMCID: PMC6685008
- DOI: 10.1098/rsif.2019.0103
Bumblebees learn foraging routes through exploitation-exploration cycles
Abstract
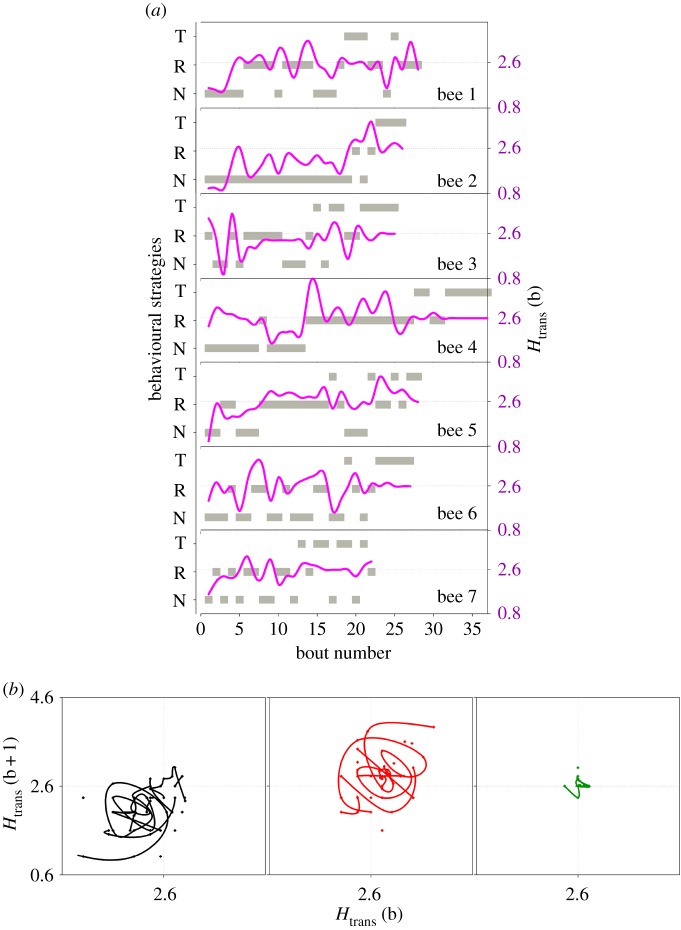
How animals explore and acquire knowledge from the environment is a key question in movement ecology. For pollinators that feed on multiple small replenishing nectar resources, the challenge is to learn efficient foraging routes while dynamically acquiring spatial information about new resource locations. Here, we use the behavioural mapping t-Stochastic Neighbouring Embedding algorithm and Shannon entropy to statistically analyse previously published sampling patterns of bumblebees feeding on artificial flowers in the field. We show that bumblebees modulate foraging excursions into distinctive behavioural strategies, characterizing the trade-off dynamics between (i) visiting and exploiting flowers close to the nest, (ii) searching for new routes and resources, and (iii) exploiting learned flower visitation sequences. Experienced bees combine these behavioural strategies even after they find an optimal route minimizing travel distances between flowers. This behavioural variability may help balancing energy costs-benefits and facilitate rapid adaptation to changing environments and the integration of more profitable resources in their routes.
Keywords: bumblebees; exploration–exploitation trade-off; movement ecology; t-Stochastic Neighbouring Embedding; trapline foraging.
Conflict of interest statement
We declare we have no competing interests.
Figures



References
-
- McNamara JM, Green RF, Olsson O. 2006. Bayes' theorem and its application in animal behaviour. Oikos 112, 243–251. (10.1111/j.0030-1299.2006.14228.x) - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources