

Differential UVR8 Signal across the Stem Controls UV-B-Induced Inflorescence Phototropism
- PMID: 31289115
- PMCID: PMC6751110
- DOI: 10.1105/tpc.18.00929
Differential UVR8 Signal across the Stem Controls UV-B-Induced Inflorescence Phototropism
Abstract
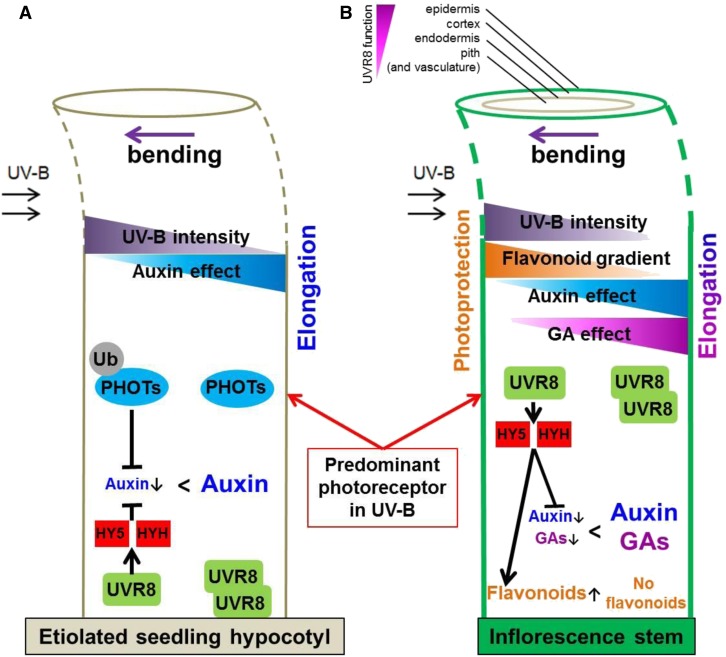
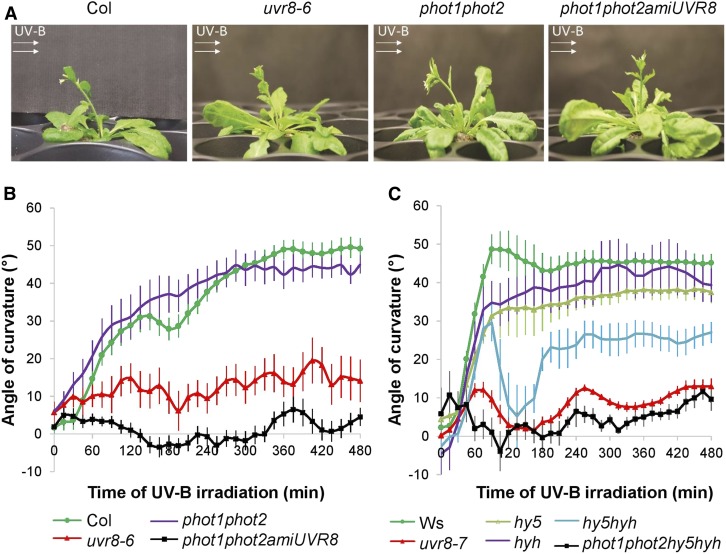
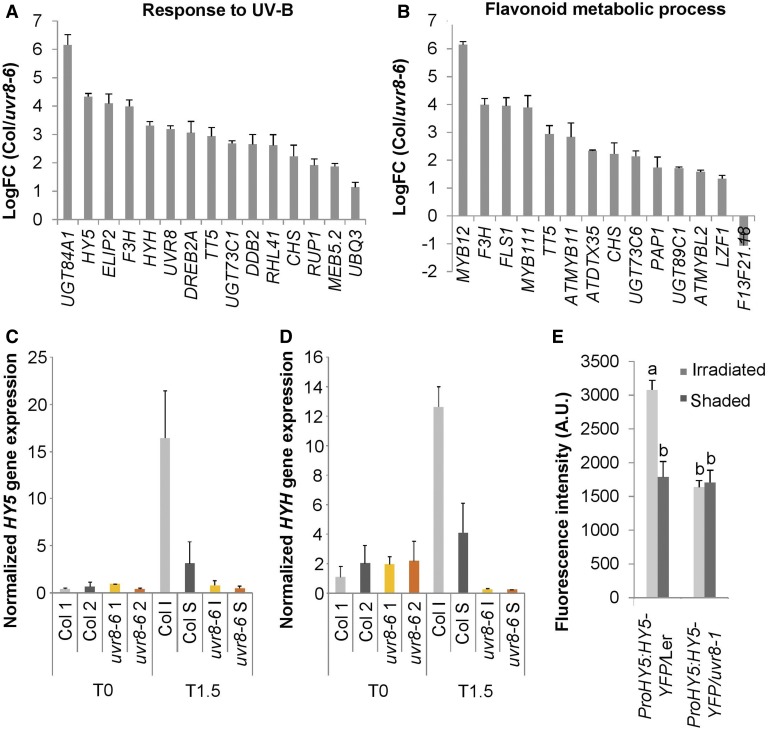
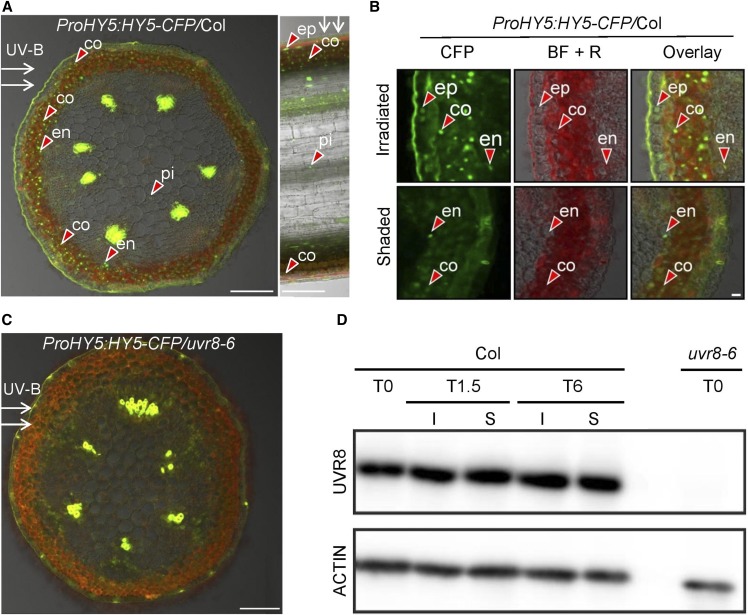
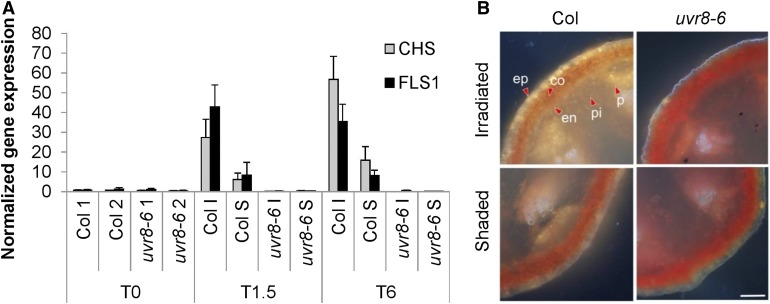
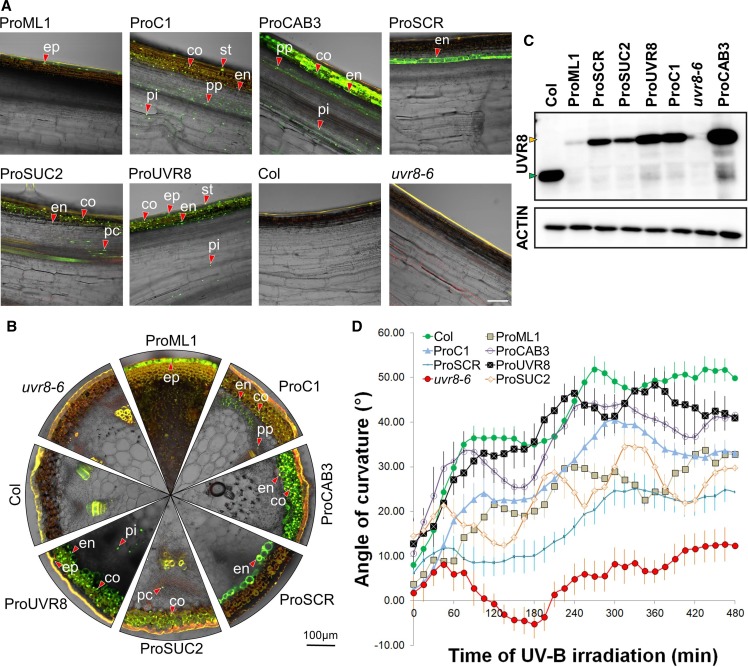
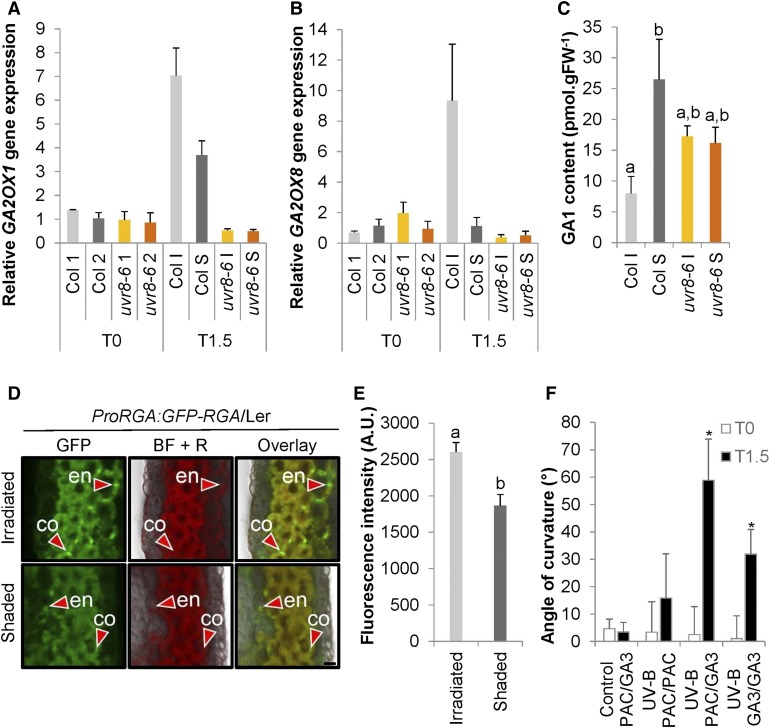
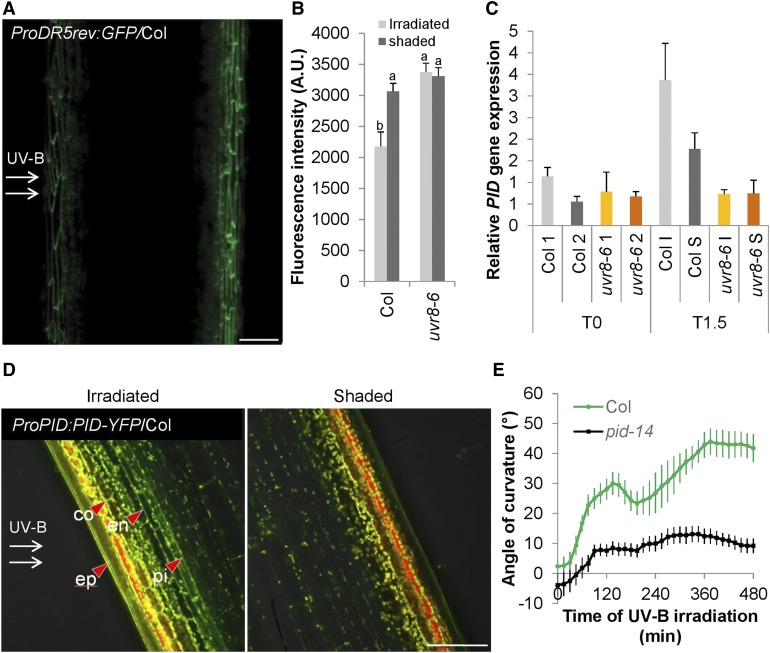
In the course of evolution, plants have developed mechanisms that orient their organs toward the incoming light. At the seedling stage, positive phototropism is mainly regulated by phototropin photoreceptors in blue and UV wavelengths. Contrasting with this, we report that UV RESISTANCE LOCUS8 (UVR8) serves as the predominant photoreceptor of UV-B-induced phototropic responses in Arabidopsis (Arabidopsis thaliana) inflorescence stems. We examined the molecular mechanisms underlying this response and our findings support the Blaauw theory (Blaauw, 1919), suggesting rapid differential growth through unilateral photomorphogenic growth inhibition. UVR8-dependent UV-B light perception occurs mainly in the epidermis and cortex, but deeper tissues such as endodermis can also contribute. Within stems, a spatial difference of UVR8 signal causes a transcript and protein increase of transcription factors ELONGATED HYPOCOTYL5 (HY5) and its homolog HY5 HOMOLOG at the UV-B-exposed side. The irradiated side shows (1) strong activation of flavonoid synthesis genes and flavonoid accumulation; (2) increased gibberellin (GA)2-oxidase expression, diminished GA1 levels, and accumulation of the DELLA protein REPRESSOR OF GA1; and (3) increased expression of the auxin transport regulator PINOID, contributing to diminished auxin signaling. Together, the data suggest a mechanism of phototropin-independent inflorescence phototropism through multiple, locally UVR8-regulated hormone pathways.
© 2019 American Society of Plant Biologists. All rights reserved.
Figures

References
-
- Atamian H.S., Creux N.M., Brown E.A., Garner A.G., Blackman B.K., Harmer S.L. (2016). Circadian regulation of sunflower heliotropism, floral orientation, and pollinator visits. Science 353: 587–590. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
