

Defining a Spinal Microcircuit that Gates Myelinated Afferent Input: Implications for Tactile Allodynia
- PMID: 31291586
- PMCID: PMC6635381
- DOI: 10.1016/j.celrep.2019.06.040
Defining a Spinal Microcircuit that Gates Myelinated Afferent Input: Implications for Tactile Allodynia
Abstract
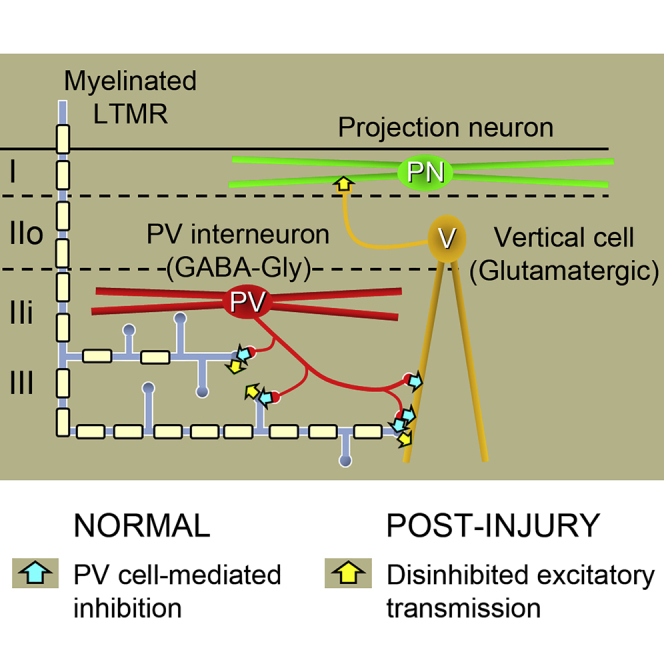
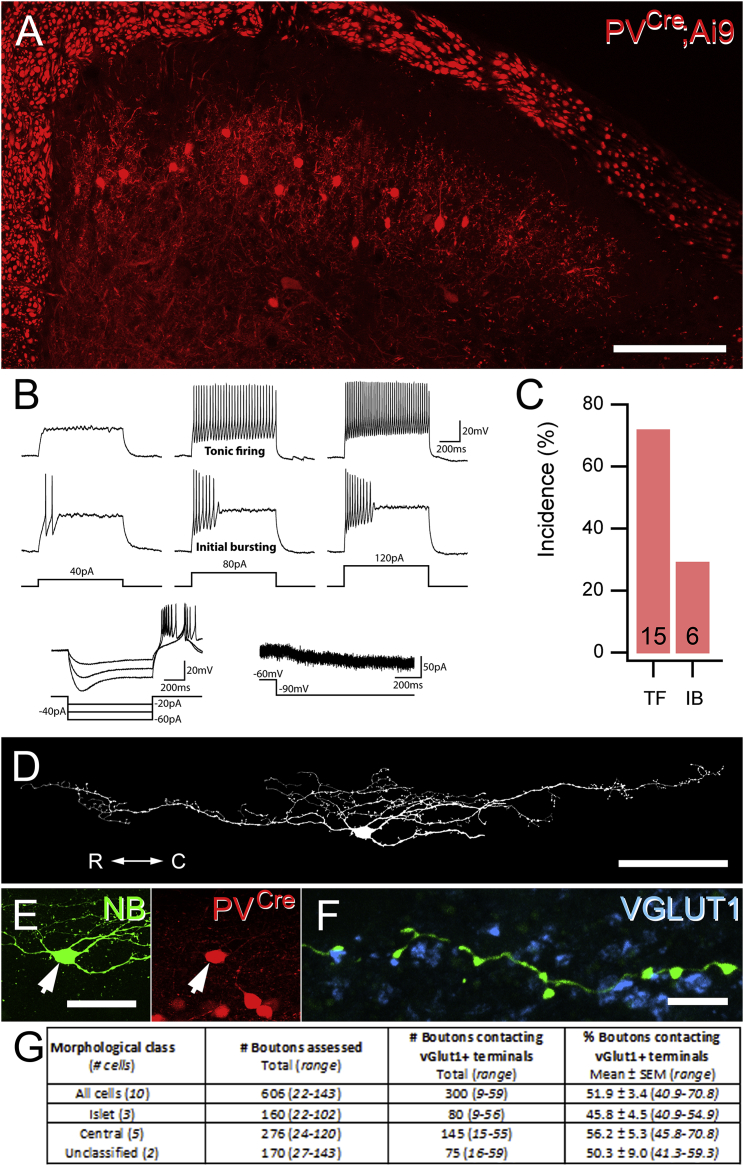
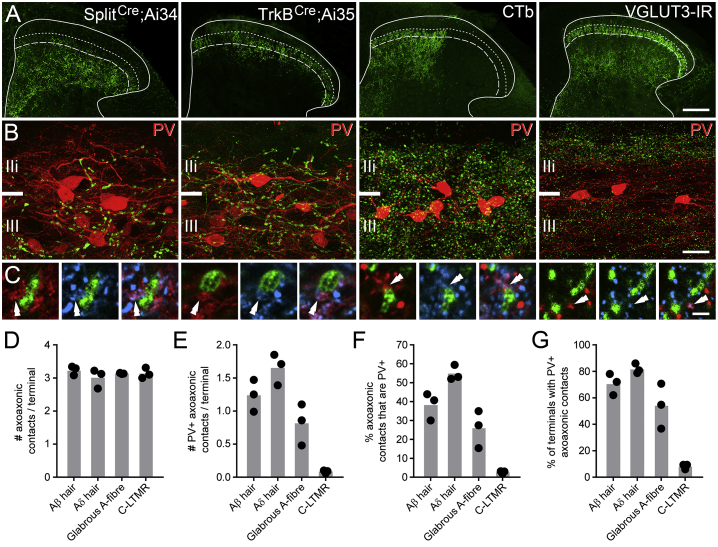
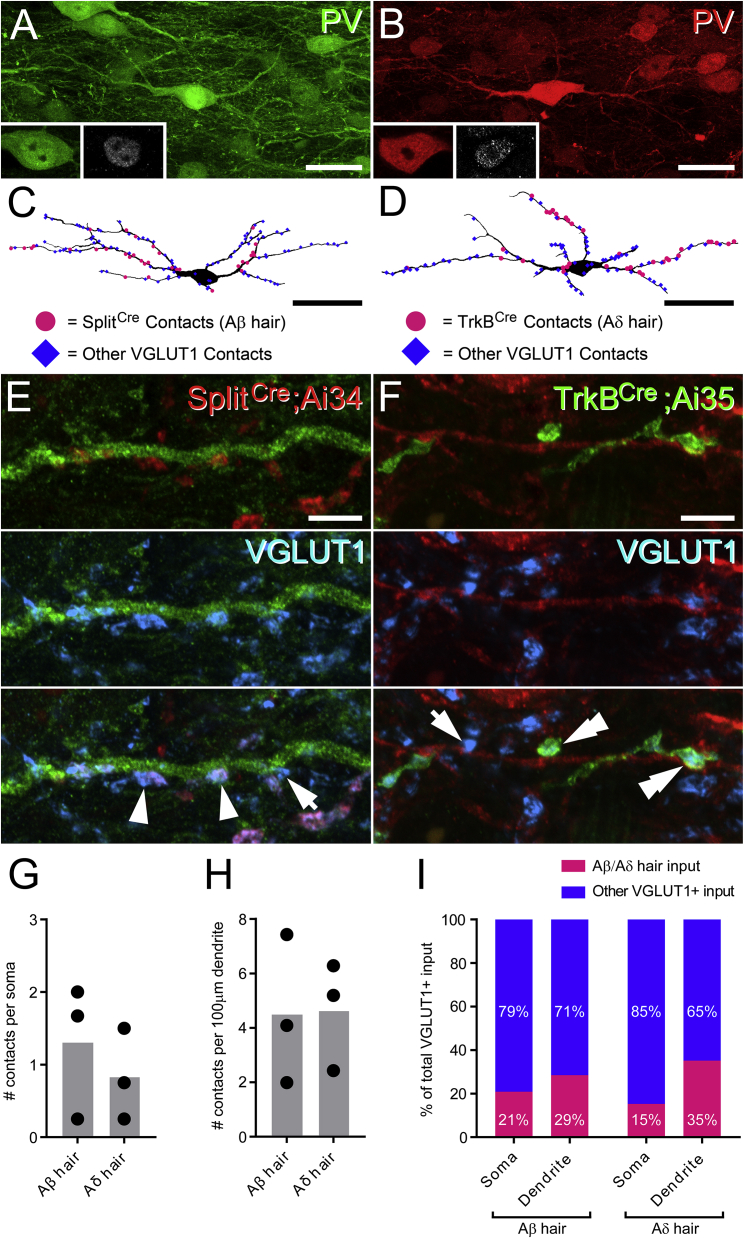
Chronic pain presents a major unmet clinical problem. The development of more effective treatments is hindered by our limited understanding of the neuronal circuits underlying sensory perception. Here, we show that parvalbumin (PV)-expressing dorsal horn interneurons modulate the passage of sensory information conveyed by low-threshold mechanoreceptors (LTMRs) directly via presynaptic inhibition and also gate the polysynaptic relay of LTMR input to pain circuits by inhibiting lamina II excitatory interneurons whose axons project into lamina I. We show changes in the functional properties of these PV interneurons following peripheral nerve injury and that silencing these cells unmasks a circuit that allows innocuous touch inputs to activate pain circuits by increasing network activity in laminae I-IV. Such changes are likely to result in the development of tactile allodynia and could be targeted for more effective treatment of mechanical pain.
Keywords: LTMRs; allodynia; dorsal horn; interneurons; parvalbumin; presynaptic inhibition; touch.
Copyright © 2019 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Ahmadi S., Lippross S., Neuhuber W.L., Zeilhofer H.U. PGE(2) selectively blocks inhibitory glycinergic neurotransmission onto rat superficial dorsal horn neurons. Nat. Neurosci. 2002;5:34–40. - PubMed
-
- Baba H., Ji R.-R., Kohno T., Moore K.A., Ataka T., Wakai A., Okamoto M., Woolf C.J. Removal of GABAergic inhibition facilitates polysynaptic A fiber-mediated excitatory transmission to the superficial spinal dorsal horn. Mol. Cell. Neurosci. 2003;24:818–830. - PubMed
-
- Bailey A.L., Ribeiro-da-Silva A. Transient loss of terminals from non-peptidergic nociceptive fibers in the substantia gelatinosa of spinal cord following chronic constriction injury of the sciatic nerve. Neuroscience. 2006;138:675–690. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
