

Spotlight on the Energy Harvest of Electroactive Microorganisms: The Impact of the Applied Anode Potential
- PMID: 31293531
- PMCID: PMC6606774
- DOI: 10.3389/fmicb.2019.01352
Spotlight on the Energy Harvest of Electroactive Microorganisms: The Impact of the Applied Anode Potential
Erratum in
-
Corrigendum: Spotlight on the Energy Harvest of Electroactive Microorganisms: The Impact of the Applied Anode Potential.Front Microbiol. 2019 Nov 28;10:2744. doi: 10.3389/fmicb.2019.02744. eCollection 2019. Front Microbiol. 2019. PMID: 31839792 Free PMC article.
Abstract
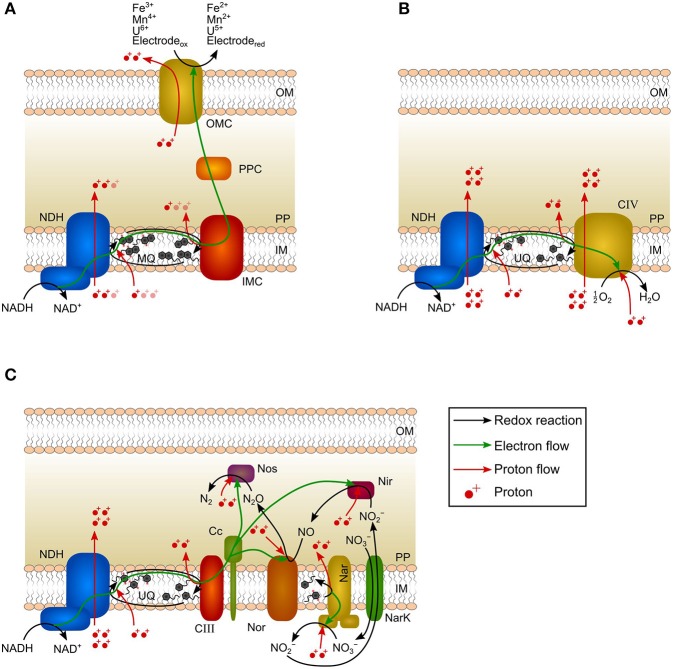
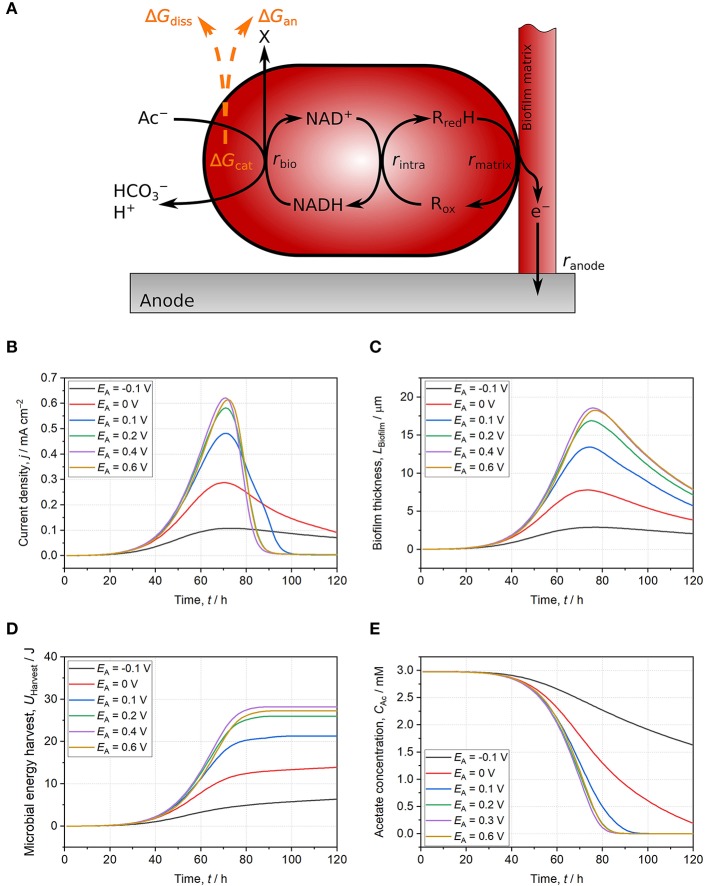
Electroactive microorganisms (EAM) harvest energy by reducing insoluble terminal electron acceptors (TEA) including electrodes via extracellular electron transfer (EET). Therefore, compared to microorganisms respiring soluble TEA, an adapted approach is required for thermodynamic analyses. In EAM, the thermodynamic frame (i.e., maximum available energy) is restricted as only a share of the energy difference between electron donor and TEA is exploited via the electron-transport chain to generate proton-motive force being subsequently utilized for ATP synthesis. However, according to a common misconception, the anode potential is suggested to co-determine the thermodynamic frame of EAM. By comparing the model organism Geobacter spp. and microorganisms respiring soluble TEA, we reason that a considerable part of the electron-transport chain of EAM performing direct EET does not contribute to the build-up of proton-motive force and thus, the anode potential does not co-determine the thermodynamic frame. Furthermore, using a modeling platform demonstrates that the influence of anode potential on energy harvest is solely a kinetic effect. When facing low anode potentials, NADH is accumulating due to a slow direct EET rate leading to a restricted exploitation of the thermodynamic frame. For anode potentials ≥ 0.2 V (vs. SHE), EET kinetics, NAD+/NADH ratio as well as exploitation of the thermodynamic frame are maximized, and a further potential increase does not result in higher energy harvest. Considering the limited influence of the anode potential on energy harvest of EAM is a prerequisite to improve thermodynamic analyses, microbial resource mining, and to transfer microbial electrochemical technologies (MET) into practice.
Keywords: electroactive microorganisms; electron-transport chain; extracellular electron transfer; microbial energy harvest; microbial thermodynamics; modeling.
Figures
References
LinkOut - more resources
Full Text Sources
Miscellaneous