

Sequential Changes in the Host Gut Microbiota During Infection With the Intestinal Parasitic Nematode Strongyloides venezuelensis
- PMID: 31293983
- PMCID: PMC6604662
- DOI: 10.3389/fcimb.2019.00217
Sequential Changes in the Host Gut Microbiota During Infection With the Intestinal Parasitic Nematode Strongyloides venezuelensis
Abstract
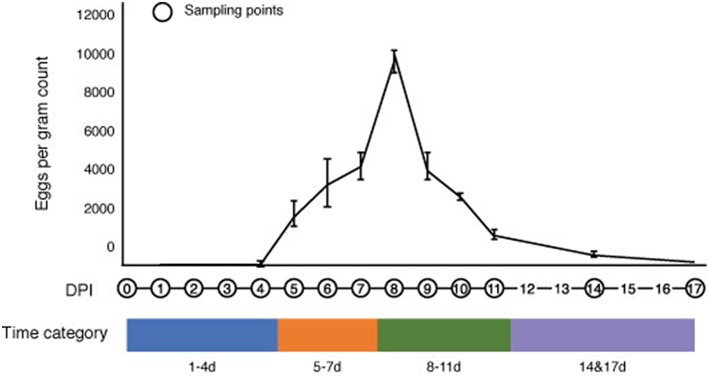
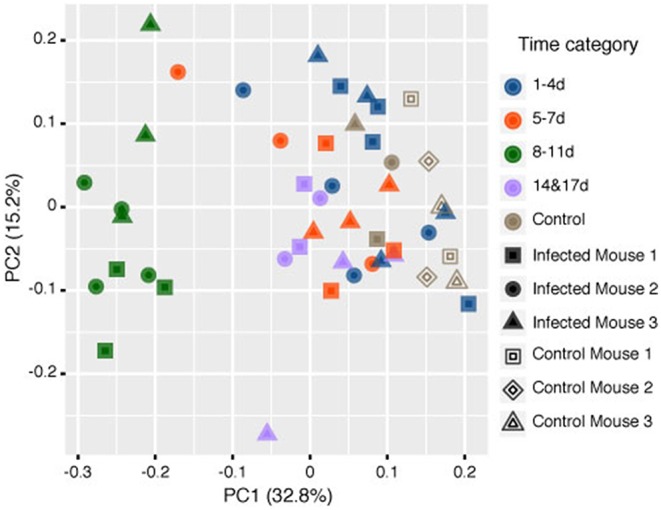
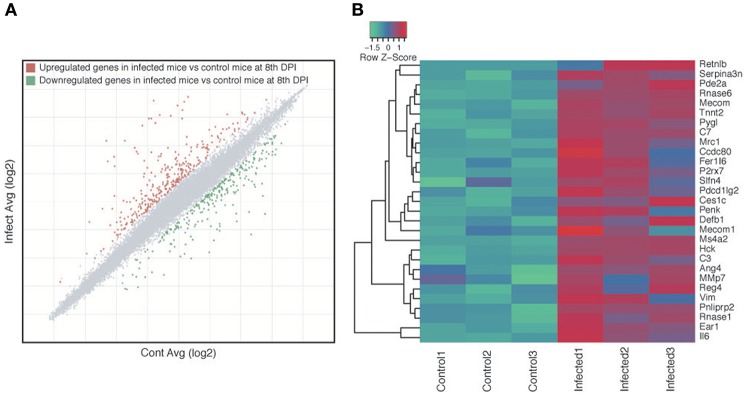
Soil-transmitted helminths (STHs) are medically important parasites that infect 1. 5 billion humans globally, causing a substantial disease burden. These parasites infect the gastrointestinal tract (GIT) of their host where they co-exist and interact with the host gut bacterial flora, leading to the coevolution of the parasites, microbiota, and host organisms. However, little is known about how these interactions change through time with the progression of infection. Strongyloidiasis is a human parasitic disease caused by the nematode Strongyloides stercoralis infecting 30-100 million people. In this study, we used a closely related rodent parasite Strongyloides venezuelensis and mice as a model of gastrointestinal parasite infection. We conducted a time-course experiment to examine changes in the fecal microbiota from the start of infection to parasite clearance. We found that bacterial taxa in the host intestinal microbiota changed significantly as the infection progressed, with an increase in the genera Bacteroides and Candidatus Arthromitus, and a decrease in Prevotella and Rikenellaceae. However, the microbiota recovered to the pre-infective state after parasite clearance from the host, suggesting that these perturbations are reversible. Microarray analysis revealed that this microbiota transition is likely to correspond with the host immune response. These findings give us an insight into the dynamics of parasite-microbiota interactions in the host gut during parasite infection.
Keywords: Candidatus Arthromitus; Strongyloides; host–parasite interaction; immune reaction; microbiome.
Figures



References
-
- Afrin T., Kounosu A., Billah M.-M., Murase K., Kikuchi T. (2018). Evaluation of magnetic cellulose bead-based DNA extraction from faecal materials for high-throughput bacterial community analyses. Appl. Entomol. Zool. 53, 281–286. 10.1007/s13355-018-0551-2 - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources