

Human Trophoblast Differentiation Is Associated With Profound Gene Regulatory and Epigenetic Changes
- PMID: 31294776
- PMCID: PMC6821221
- DOI: 10.1210/en.2019-00144
Human Trophoblast Differentiation Is Associated With Profound Gene Regulatory and Epigenetic Changes
Abstract
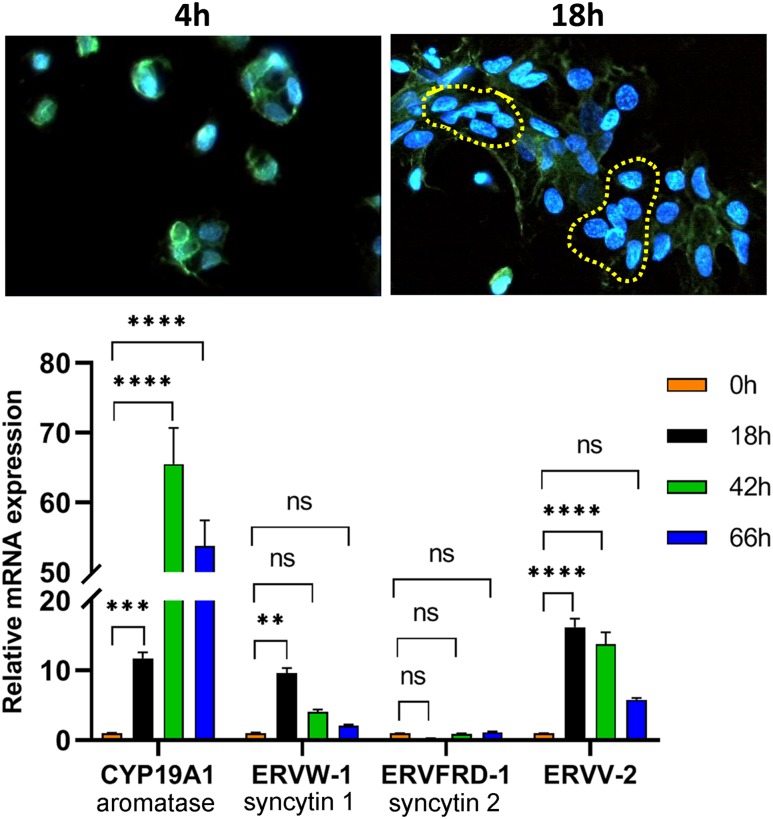
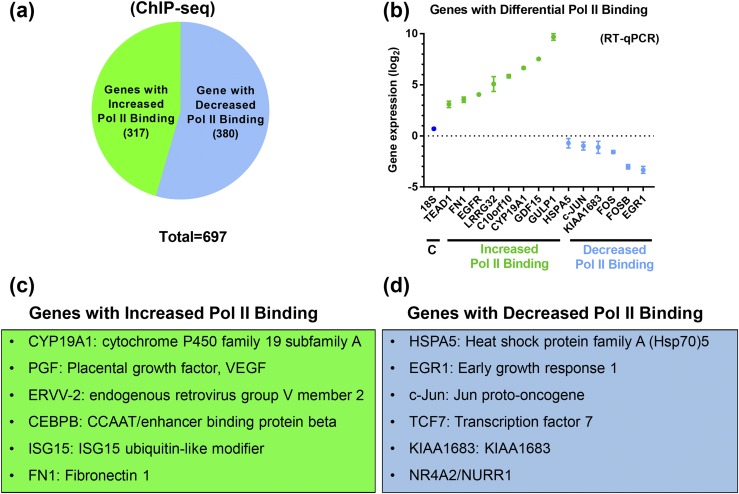
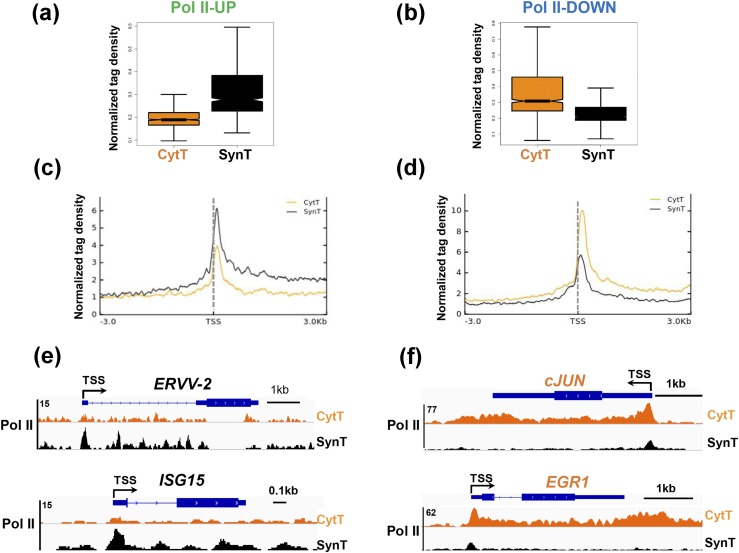
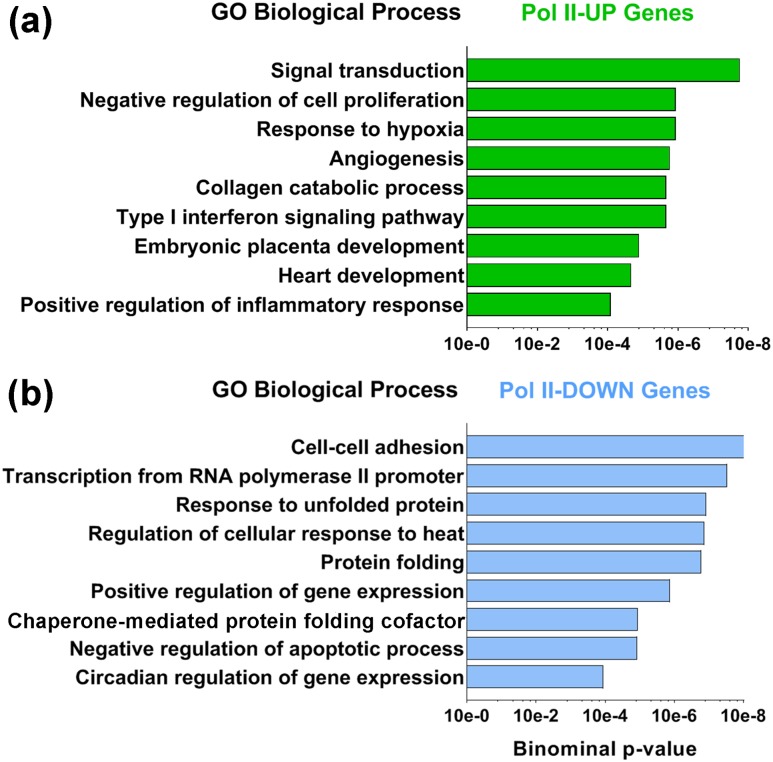
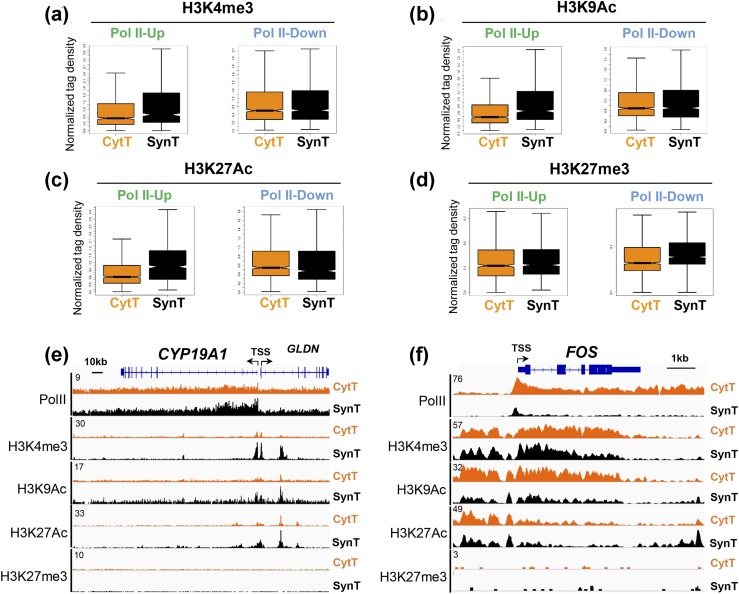
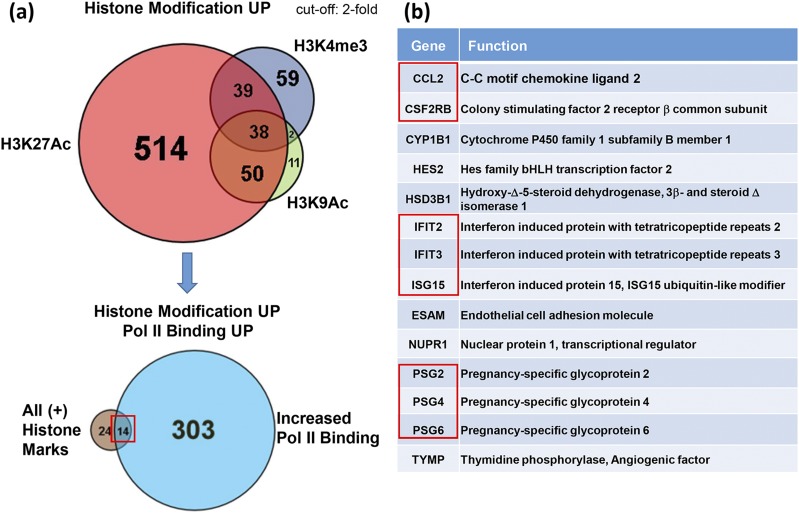
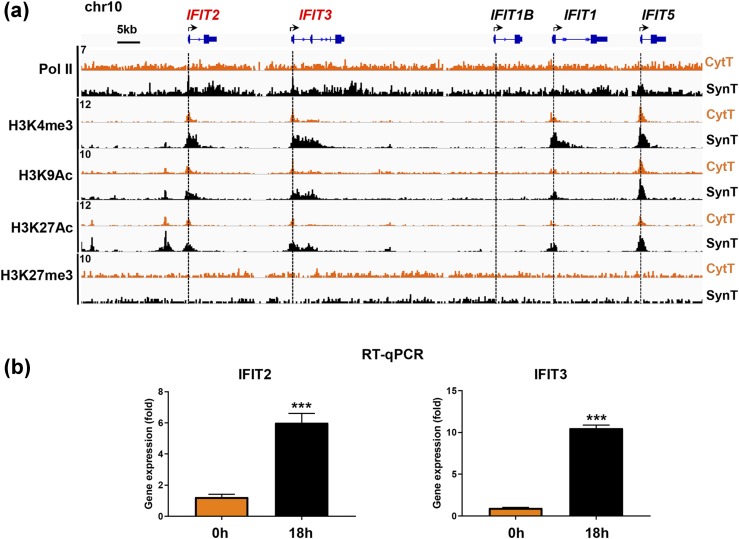
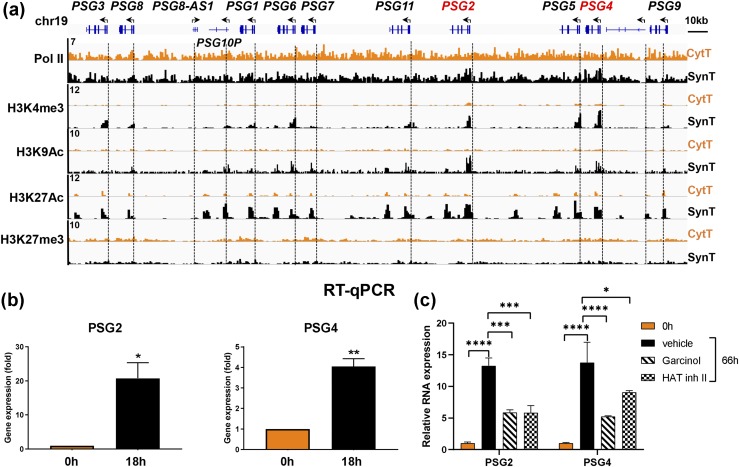
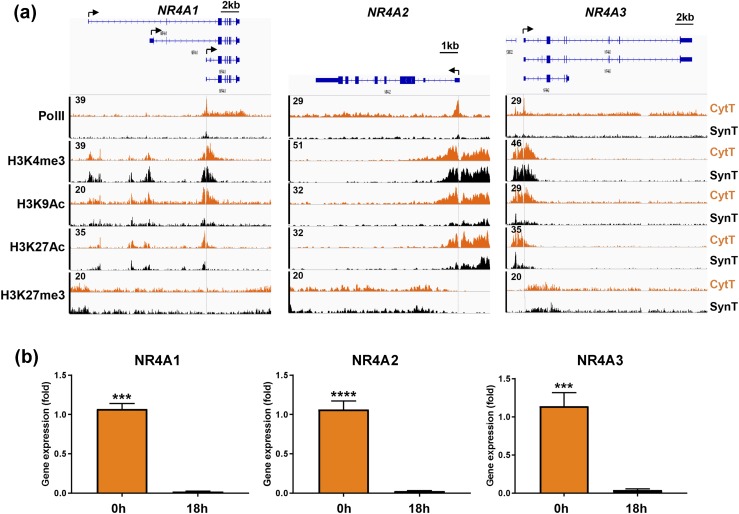
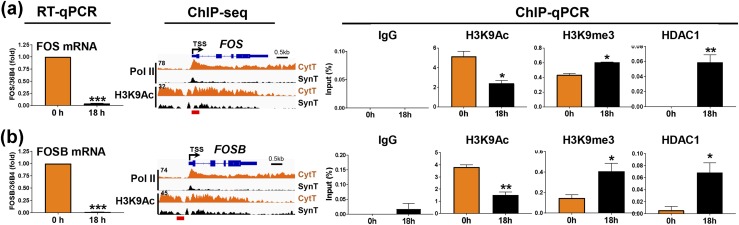
Defective placental implantation and vascularization with accompanying hypoxia contribute to preeclampsia (PE), a leading cause of maternal and neonatal morbidity and mortality. Genetic and epigenetic mechanisms underlying differentiation of proliferative cytotrophoblasts (CytTs) to multinucleated syncytiotrophoblast (SynT) are incompletely defined. The SynT performs key functions in nutrient and gas exchange, hormone production, and protection of the fetus from rejection by the maternal immune system. In this study, we used chromatin immunoprecipitation sequencing of midgestation human trophoblasts before CytT and after SynT differentiation in primary culture to analyze changes in binding of RNA polymerase II (Pol II) and of active and repressive histone marks during SynT differentiation. Our findings reveal that increased Pol II binding to promoters of a subset of genes during trophoblast differentiation was closely correlated with active histone marks. This gene set was enriched in those controlling immune response and immune modulation, including interferon-induced tetratricopeptide repeat and placenta-specific glycoprotein gene family members. By contrast, genes downregulated during SynT differentiation included proinflammatory transcription factors ERG1, cFOS, and cJUN, as well as members of the NR4A orphan nuclear receptor subfamily, NUR77, NURR1, and NOR1. Downregulation of proinflammatory transcription factors upon SynT differentiation was associated with decreased promoter enrichment of endogenous H3K27Ac and H3K9Ac and enhanced binding of H3K9me3 and histone deacetylase 1. However, promoter enrichment of H3K27me3 was low in both CytT and SynT and was not altered with changes in gene expression. These findings provide important insight into mechanisms underlying human trophoblast differentiation and may identify therapeutic targets for placental disorders, such as PE.
Copyright © 2019 Endocrine Society.
Conflict of interest statement
All data generated or analyzed during this study are included in this published article or in the data repositories listed in References.
Figures
References
-
- Everett RB, MacDonald PC. Endocrinology of the placenta. Annu Rev Med. 1979;30(1):473–488. - PubMed
-
- McMaster MT, Zhou Y, Fisher SJ. Abnormal placentation and the syndrome of preeclampsia. Semin Nephrol. 2004;24(6):540–547. - PubMed
-
- Arck PC, Hecher K. Fetomaternal immune cross-talk and its consequences for maternal and offspring’s health. Nat Med. 2013;19(5):548–556. - PubMed
-
- PrabhuDas M, Bonney E, Caron K, Dey S, Erlebacher A, Fazleabas A, Fisher S, Golos T, Matzuk M, McCune JM, Mor G, Schulz L, Soares M, Spencer T, Strominger J, Way SS, Yoshinaga K. Immune mechanisms at the maternal-fetal interface: perspectives and challenges. Nat Immunol. 2015;16(4):328–334. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
