

The plastidial pentose phosphate pathway is essential for postglobular embryo development in Arabidopsis
- PMID: 31296566
- PMCID: PMC6660741
- DOI: 10.1073/pnas.1908556116
The plastidial pentose phosphate pathway is essential for postglobular embryo development in Arabidopsis
Abstract
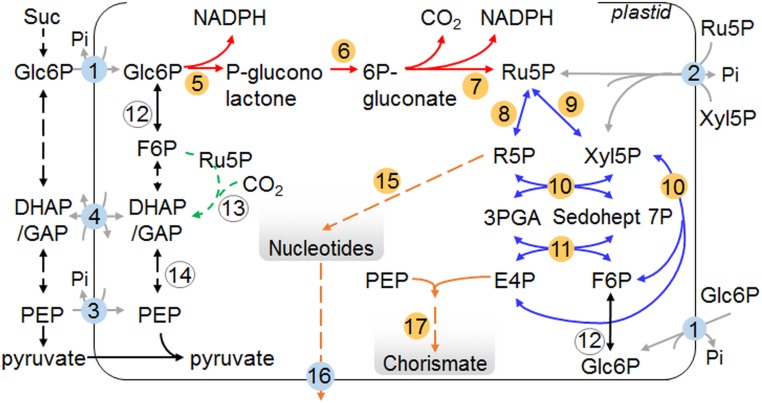
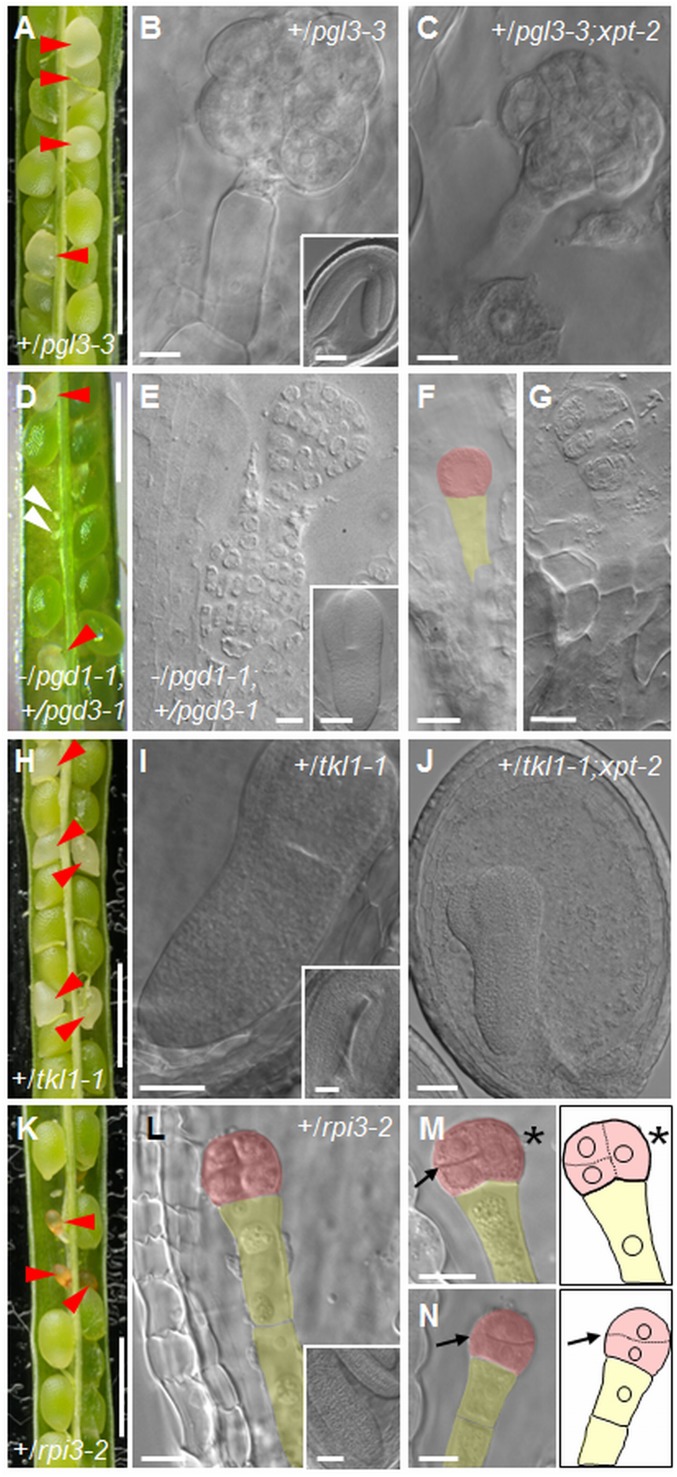
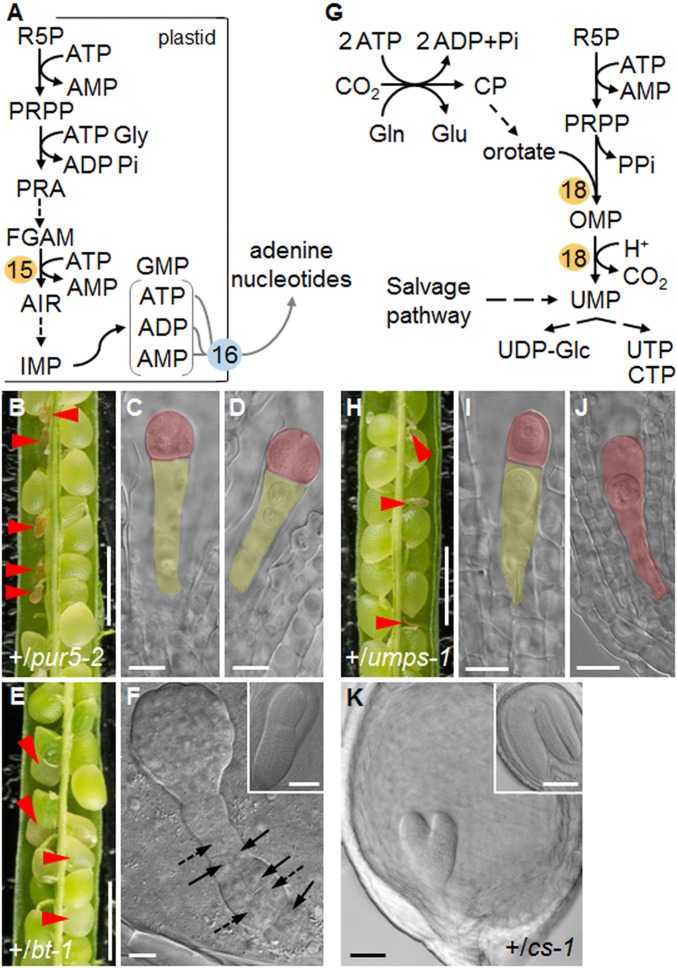
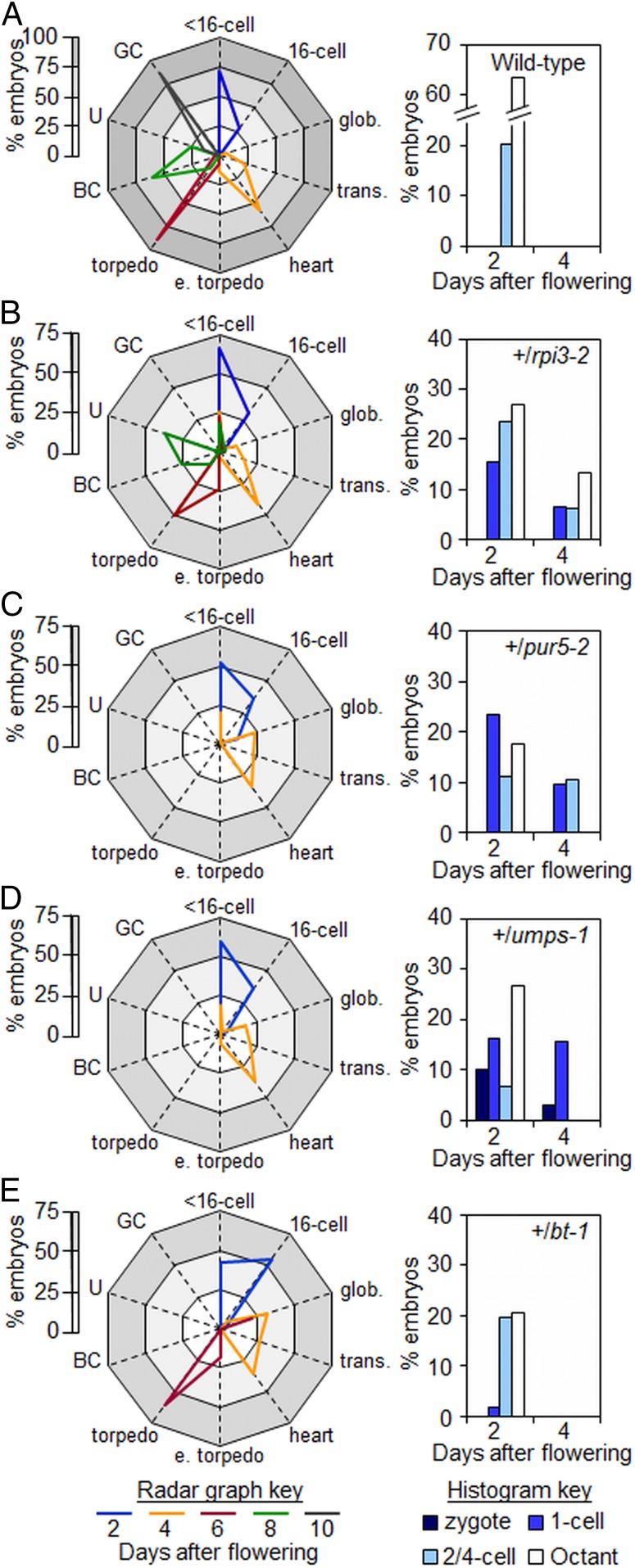
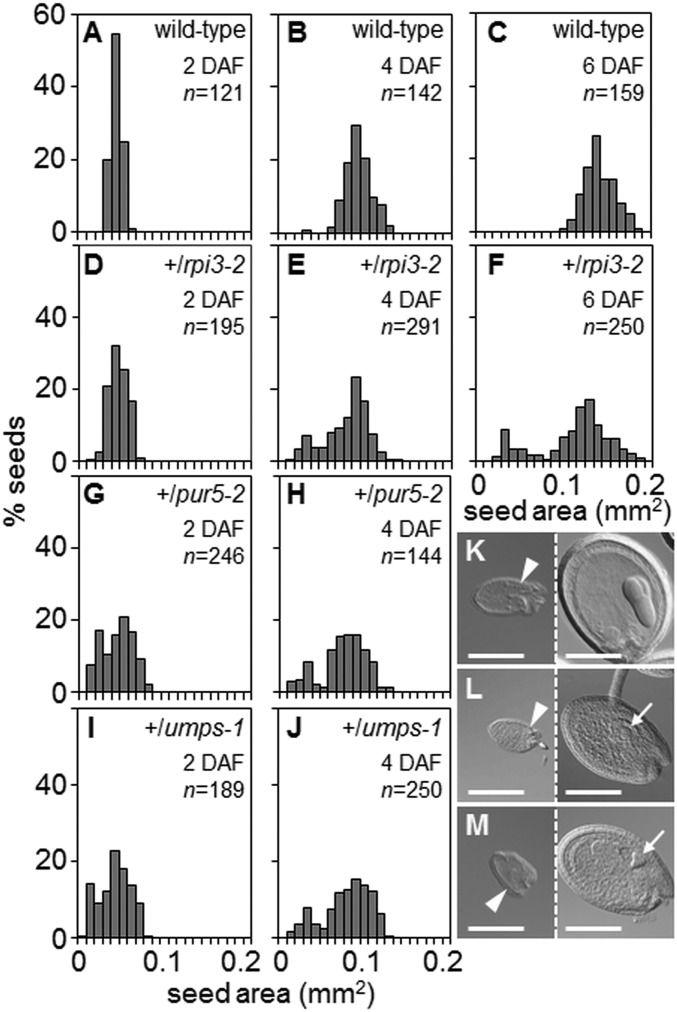
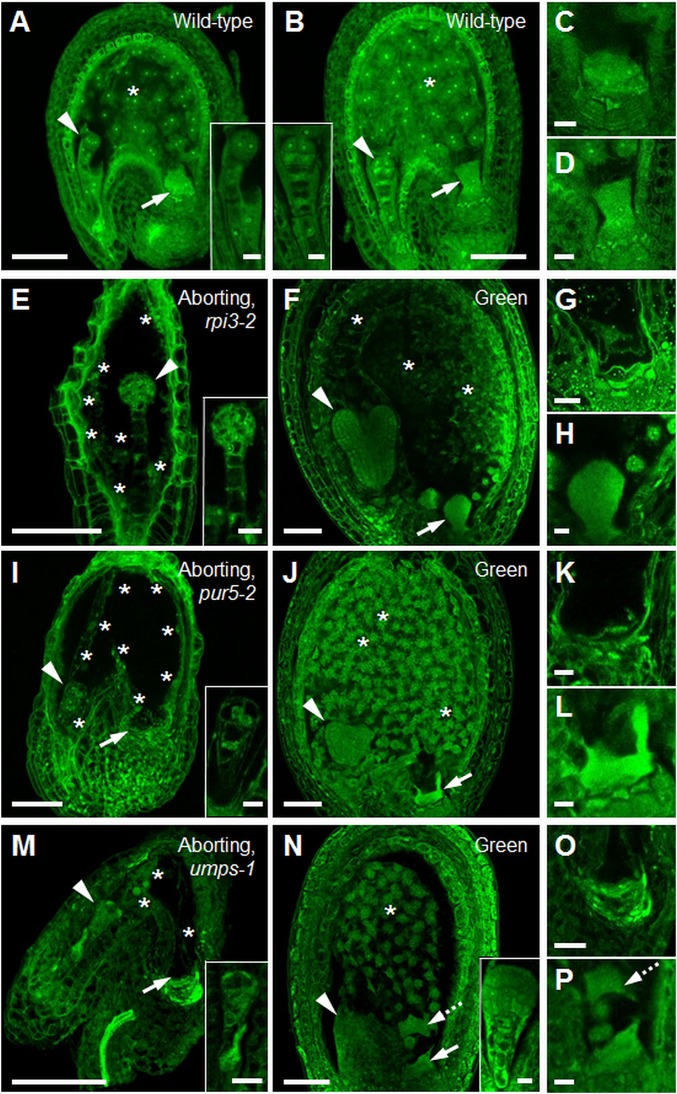
Large numbers of genes essential for embryogenesis in Arabidopsis encode enzymes of plastidial metabolism. Disruption of many of these genes results in embryo arrest at the globular stage of development. However, the cause of lethality is obscure. We examined the role of the plastidial oxidative pentose phosphate pathway (OPPP) in embryo development. In nonphotosynthetic plastids the OPPP produces reductant and metabolic intermediates for central biosynthetic processes. Embryos with defects in various steps in the oxidative part of the OPPP had cell division defects and arrested at the globular stage, revealing an absolute requirement for the production via these steps of ribulose-5-phosphate. In the nonoxidative part of the OPPP, ribulose-5-phosphate is converted to ribose-5-phosphate (R5P)-required for purine nucleotide and histidine synthesis-and subsequently to erythrose-4-phosphate, which is required for synthesis of aromatic amino acids. We show that embryo development through the globular stage specifically requires synthesis of R5P rather than erythrose-4-phosphate. Either a failure to convert ribulose-5-phosphate to R5P or a block in purine nucleotide biosynthesis beyond R5P perturbs normal patterning of the embryo, disrupts endosperm development, and causes early developmental arrest. We suggest that seed abortion in mutants unable to synthesize R5P via the oxidative part of the OPPP stems from a lack of substrate for synthesis of purine nucleotides, and hence nucleic acids. Our results show that the plastidial OPPP is essential for normal developmental progression as well as for growth in the embryo.
Keywords: Arabidopsis; embryo; nucleotide synthesis; pentose phosphate pathway; plastid.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Meinke D. W., Embryo-lethal mutants of Arabidopsis thaliana: Analysis of mutants with a wide range of lethal phases. Theor. Appl. Genet. 69, 543–552 (1985). - PubMed
-
- Mansfield S. G., Briarty L. G., Early embryogenesis in Arabidopsis thaliana. II. The developing embryo. Can. J. Bot. 69, 461–476 (1991).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
