

Large-conductance calcium-activated potassium channels mediate lipopolysaccharide-induced activation of murine microglia
- PMID: 31296663
- PMCID: PMC6721947
- DOI: 10.1074/jbc.RA118.006425
Large-conductance calcium-activated potassium channels mediate lipopolysaccharide-induced activation of murine microglia
Abstract
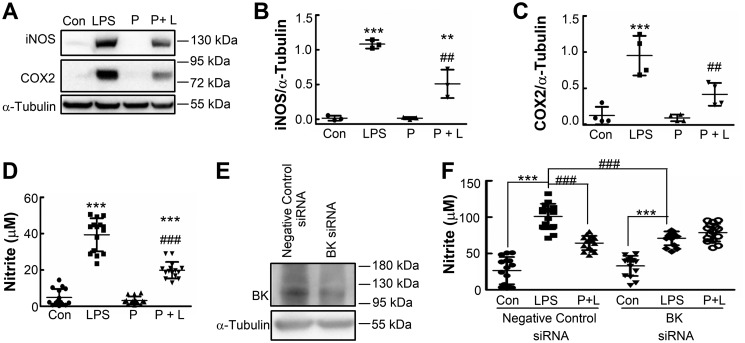
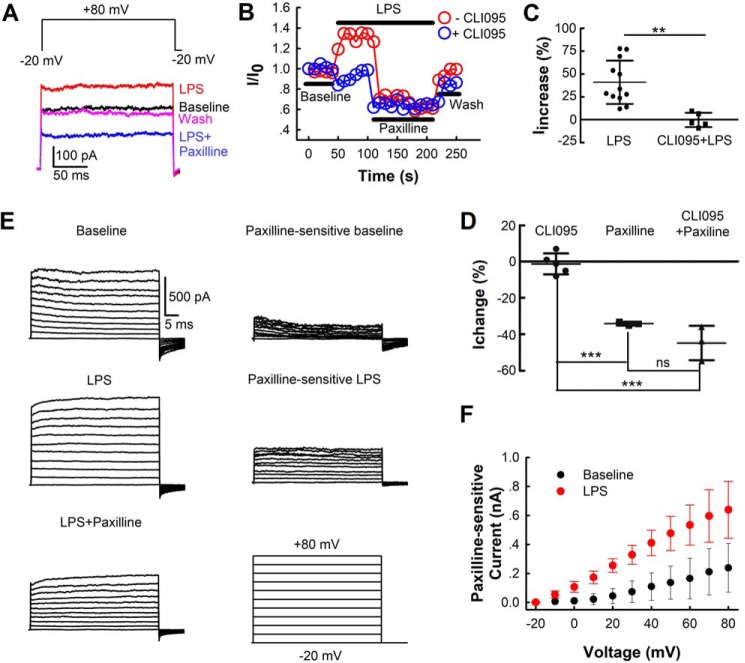
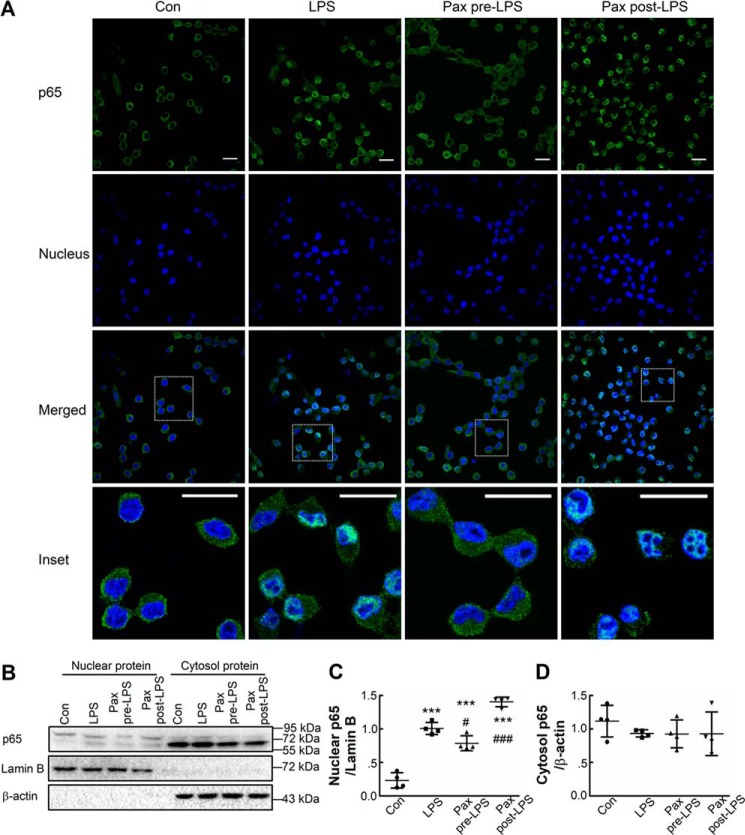
Large-conductance calcium-activated potassium (BK) channels are ubiquitously expressed in most cell types where they regulate many cellular, organ, and organismal functions. Although BK currents have been recorded specifically in activated murine and human microglia, it is not yet clear whether and how the function of this channel is related to microglia activation. Here, using patch-clamping, Griess reaction, ELISA, immunocytochemistry, and immunoblotting approaches, we show that specific inhibition of the BK channel with paxilline (10 μm) or siRNA-mediated knockdown of its expression significantly suppresses lipopolysaccharide (LPS)-induced (100 ng/ml) BV-2 and primary mouse microglial cell activation. We found that membrane BK current is activated by LPS at a very early stage through Toll-like receptor 4 (TLR4), leading to nuclear translocation of NF-κB and to production of inflammatory cytokines. Furthermore, we noted that BK channels are also expressed intracellularly, and their nuclear expression significantly increases in late stages of LPS-mediated microglia activation, possibly contributing to production of nitric oxide, tumor necrosis factor-α, and interleukin-6. Of note, a specific TLR4 inhibitor suppressed BK channel expression, whereas an NF-κB inhibitor did not. Taken together, our findings indicate that BK channels participate in both the early and the late stages of LPS-stimulated murine microglia activation involving both membrane-associated and nuclear BK channels.
Keywords: BK channel; NF-κB (NF-KB); Toll-like receptor 4 (TLR4); cytokine; innate immunity; lipopolysaccharide (LPS); microglia; neuroinflammation.
© 2019 Yang et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures




References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
