

Genomic diversity and novel genome-wide association with fruit morphology in Capsicum, from 746k polymorphic sites
- PMID: 31296904
- PMCID: PMC6624249
- DOI: 10.1038/s41598-019-46136-5
Genomic diversity and novel genome-wide association with fruit morphology in Capsicum, from 746k polymorphic sites
Abstract
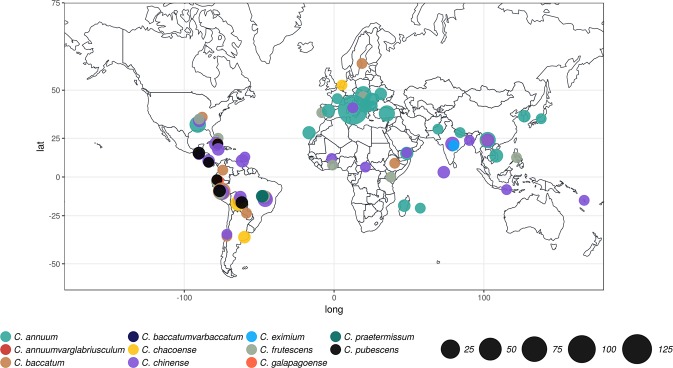
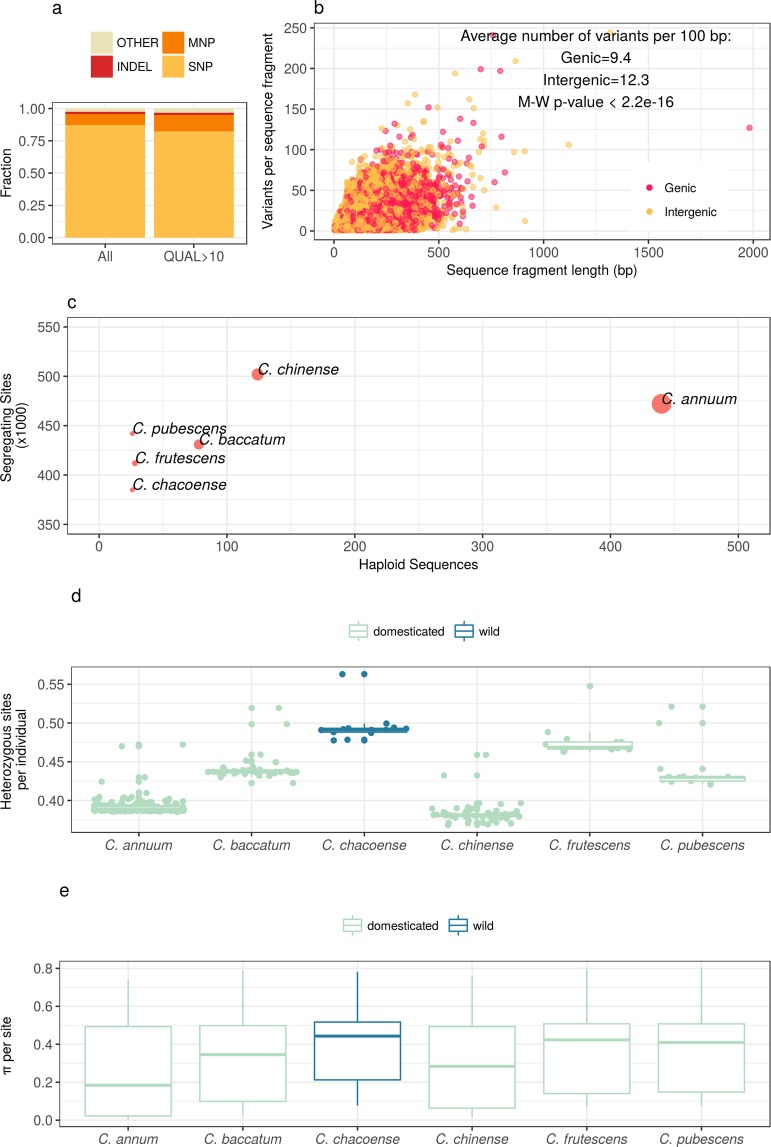
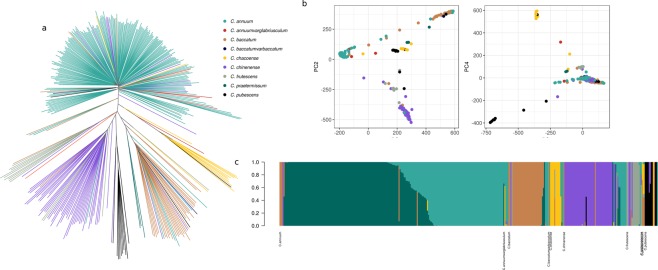
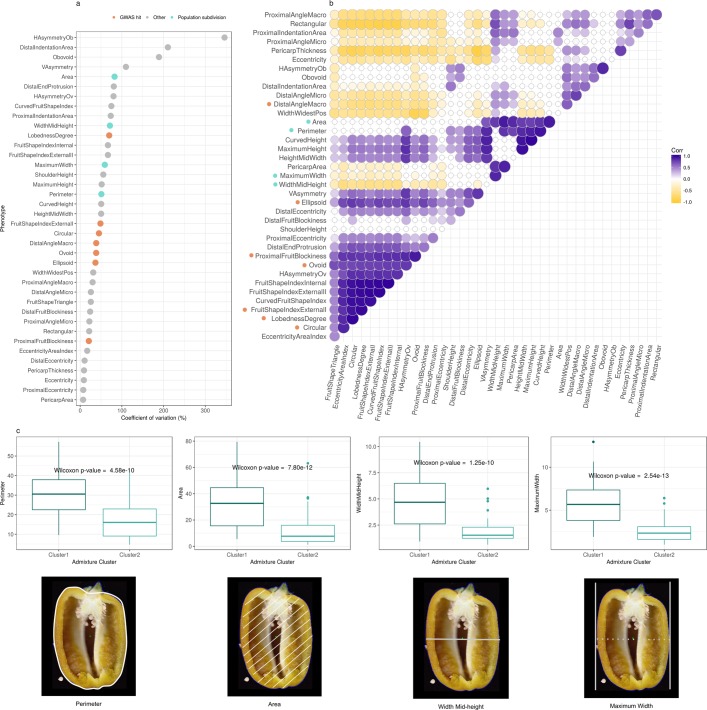
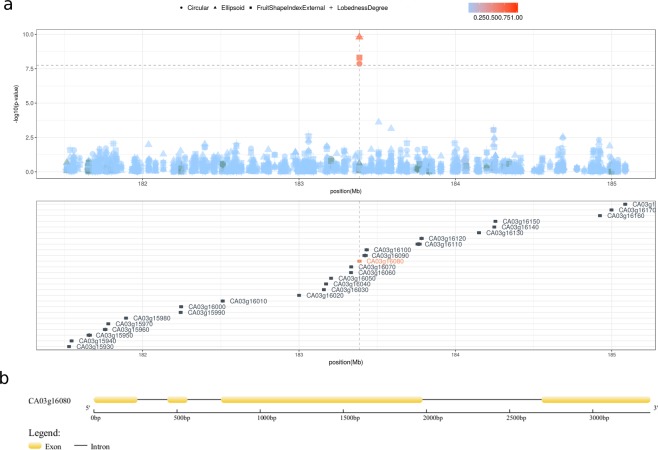
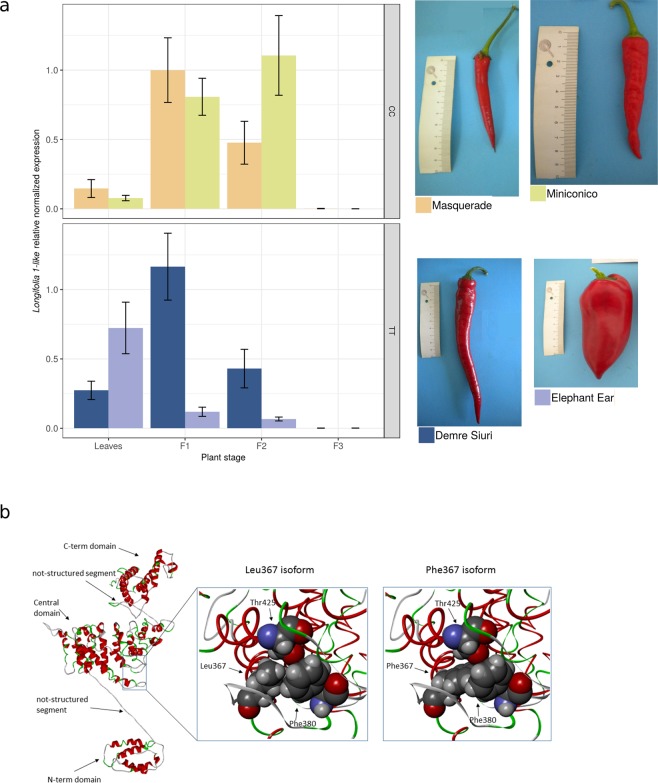
Capsicum is one of the major vegetable crops grown worldwide. Current subdivision in clades and species is based on morphological traits and coarse sets of genetic markers. Broad variability of fruits has been driven by breeding programs and has been mainly studied by linkage analysis. We discovered 746k variable sites by sequencing 1.8% of the genome in a collection of 373 accessions belonging to 11 Capsicum species from 51 countries. We describe genomic variation at population-level, confirm major subdivision in clades and species, and show that the known major subdivision of C. annuum separates large and bulky fruits from small ones. In C. annuum, we identify four novel loci associated with phenotypes determining the fruit shape, including a non-synonymous mutation in the gene Longifolia 1-like (CA03g16080). Our collection covers all the economically important species of Capsicum widely used in breeding programs and represent the widest and largest study so far in terms of the number of species and number of genetic variants analyzed. We identified a large set of markers that can be used for population genetic studies and genetic association analyses. Our results provide a comprehensive and precise perspective on genomic variability in Capsicum at population-level and suggest that future fine genetic association studies will yield useful results for breeding.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- Nicolaï M, Cantet M, Lefebvre V, Sage-Palloix A-M, Palloix A. Genotyping a large collection of pepper (Capsicum spp.) with ssr loci brings new evidence for the wild origin of cultivated C. annuum and the structuring of genetic diversity by human selection of cultivar types. Genet. Resour. Crop. Evol. 2013;60:2375–2390. doi: 10.1007/s10722-013-0006-0. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
