

Axonal Growth of Midbrain Dopamine Neurons is Modulated by the Cell Adhesion Molecule ALCAM Through Trans-Heterophilic Interactions with L1cam, Chl1, and Semaphorins
- PMID: 31300520
- PMCID: PMC6703882
- DOI: 10.1523/JNEUROSCI.0278-19.2019
Axonal Growth of Midbrain Dopamine Neurons is Modulated by the Cell Adhesion Molecule ALCAM Through Trans-Heterophilic Interactions with L1cam, Chl1, and Semaphorins
Abstract
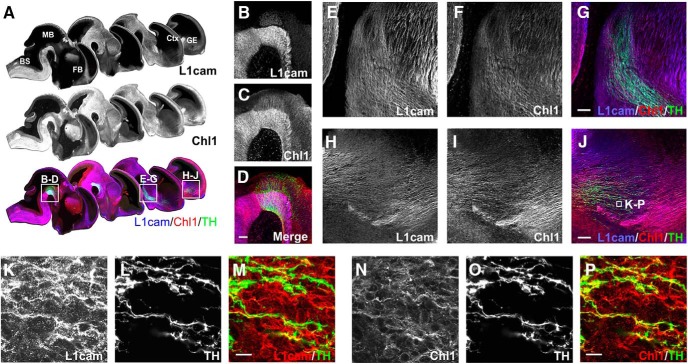
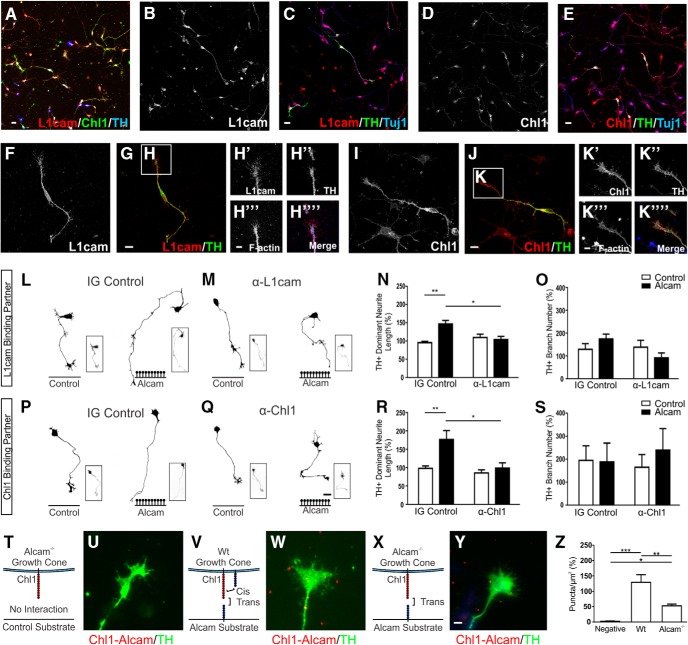
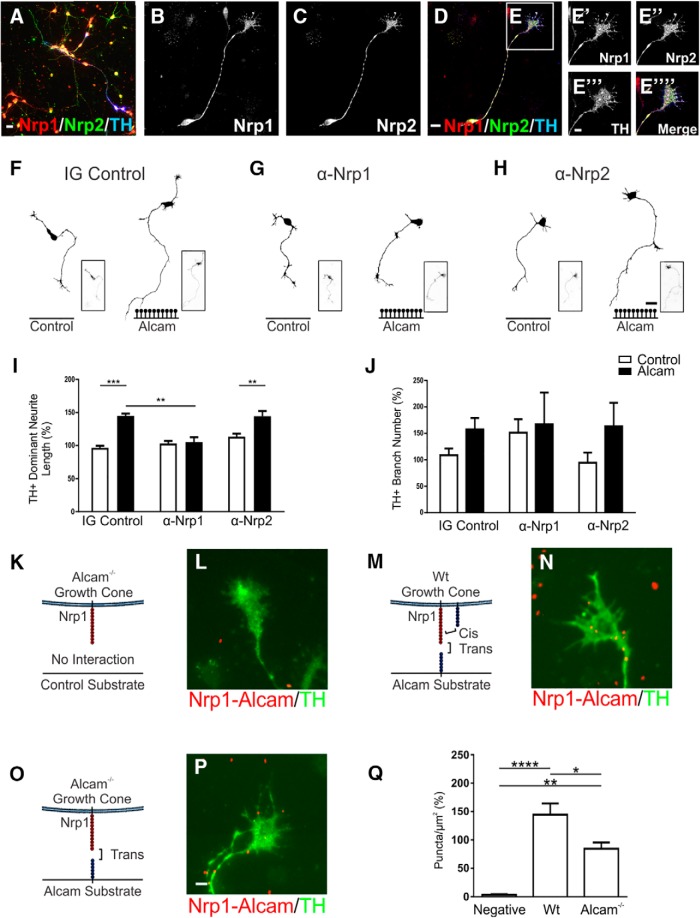
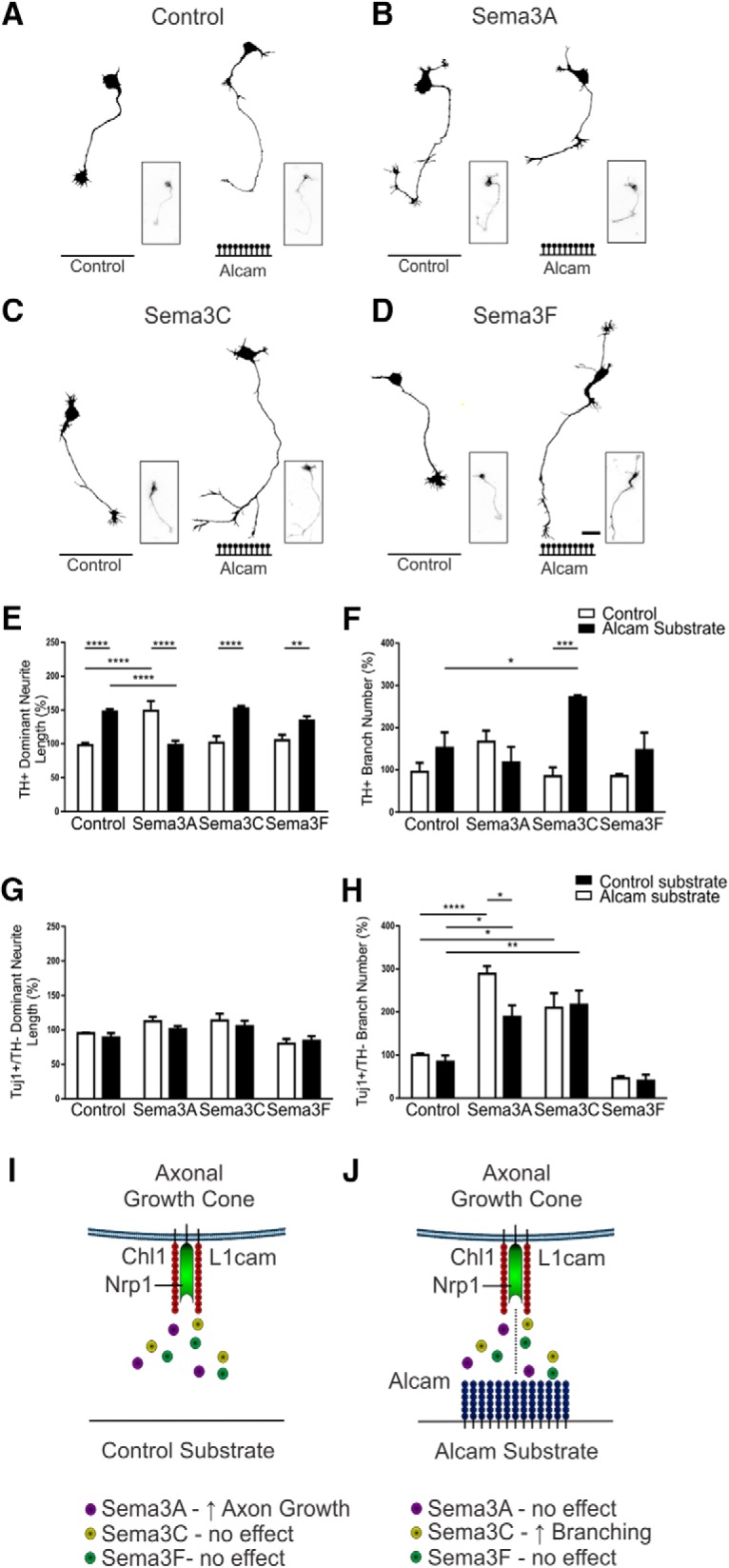
The growth of axons corresponding to different neuronal subtypes is governed by unique expression profiles of molecules on the growth cone. These molecules respond to extracellular cues either locally though cell adhesion interactions or over long distances through diffusible gradients. Here, we report that that the cell adhesion molecule ALCAM (CD166) can act as an extracellular substrate to selectively promote the growth of murine midbrain dopamine (mDA) neuron axons through a trans-heterophilic interaction with mDA-bound adhesion molecules. In mixed-sex primary midbrain cultures, the growth-promoting effect of ALCAM was abolished by neutralizing antibodies for components of the Semaphorin receptor complex Nrp1, Chl1, or L1cam. The ALCAM substrate was also found to modulate the response of mDA neurites to soluble semaphorins in a context-specific manner by abolishing the growth-promoting effect of Sema3A but inducing a branching response in the presence of Sema3C. These findings identify a previously unrecognized guidance mechanism whereby cell adhesion molecules act in trans to modulate the response of axonal growth cones to soluble gradients to selectively orchestrate the growth and guidance of mDA neurons.SIGNIFICANCE STATEMENT The mechanisms governing the axonal connectivity of midbrain dopamine (mDA) neurons during neural development have remained rather poorly understood relative to other model systems for axonal growth and guidance. Here, we report a series of novel interactions between proteins previously not identified in the context of mDA neuronal growth. Significantly, the results suggest a previously unrecognized mechanism involving the convergence in signaling between local, adhesion and long-distance, soluble cues. A better understanding of the molecules and mechanisms involved in establishment of the mDA system is important as a part of ongoing efforts to understand the consequence of conditions that may result from aberrant connectivity and also for cell replacement strategies for Parkinson's disease.
Keywords: CAMs; Nrp1; connectivity; guidance; mesencephalon.
Copyright © 2019 the authors.
Figures
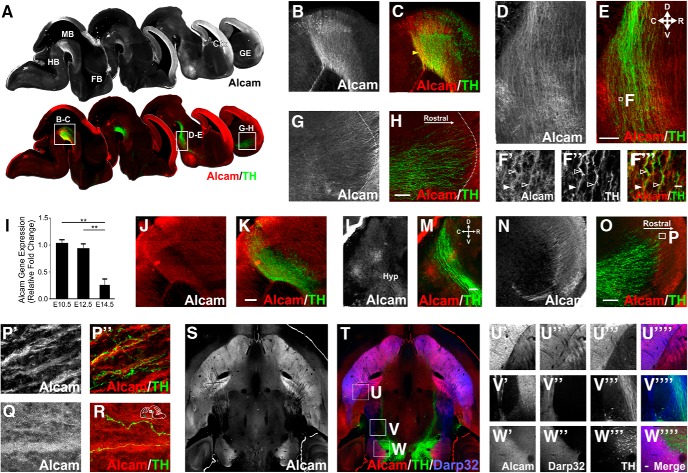
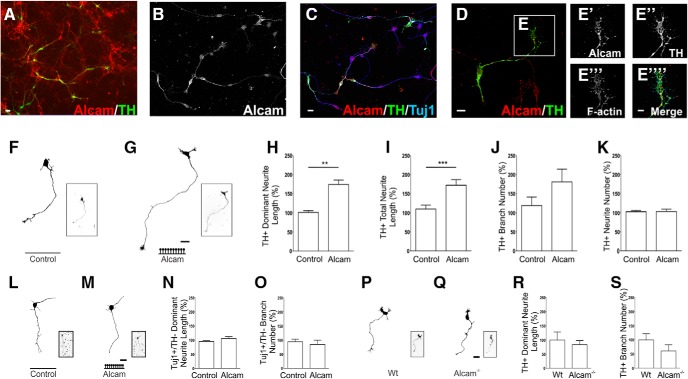
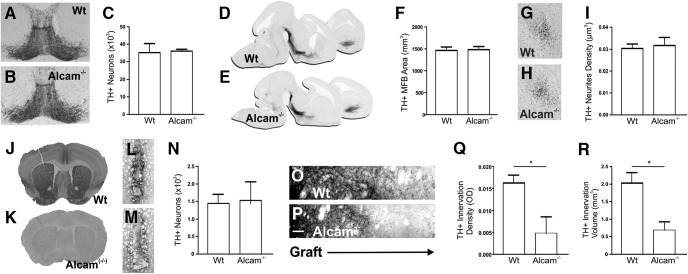
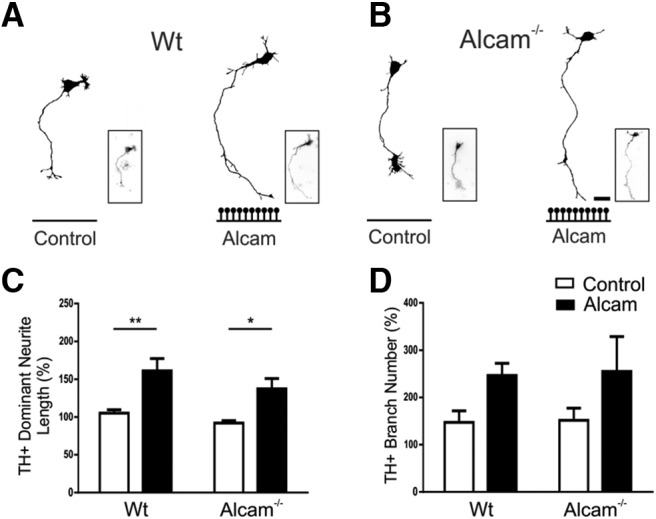
Similar articles
-
The function of neuropilin/L1 complex.Adv Exp Med Biol. 2002;515:91-102. doi: 10.1007/978-1-4615-0119-0_8. Adv Exp Med Biol. 2002. PMID: 12613546 Review.
-
Homophilic binding of the neural cell adhesion molecule CHL1 regulates development of ventral midbrain dopaminergic pathways.Sci Rep. 2017 Aug 24;7(1):9368. doi: 10.1038/s41598-017-09599-y. Sci Rep. 2017. PMID: 28839197 Free PMC article.
-
Close Homolog of L1 Regulates Dendritic Spine Density in the Mouse Cerebral Cortex Through Semaphorin 3B.J Neurosci. 2019 Aug 7;39(32):6233-6250. doi: 10.1523/JNEUROSCI.2984-18.2019. Epub 2019 Jun 10. J Neurosci. 2019. PMID: 31182634 Free PMC article.
-
FGF8 signaling regulates growth of midbrain dopaminergic axons by inducing semaphorin 3F.J Neurosci. 2009 Apr 1;29(13):4044-55. doi: 10.1523/JNEUROSCI.4794-08.2009. J Neurosci. 2009. PMID: 19339600 Free PMC article.
-
Cell signalling cascades regulating neuronal growth-promoting and inhibitory cues.Prog Neurobiol. 2001 Dec;65(6):593-608. doi: 10.1016/s0301-0082(01)00017-x. Prog Neurobiol. 2001. PMID: 11728645 Review.
Cited by
-
Alcam-a and Pdgfr-α are essential for the development of sclerotome-derived stromal cells that support hematopoiesis.Nat Commun. 2023 Mar 1;14(1):1171. doi: 10.1038/s41467-023-36612-y. Nat Commun. 2023. PMID: 36859431 Free PMC article.
-
Extracellular Matrix Biomimetic Hydrogels, Encapsulated with Stromal Cell-Derived Factor 1, Improve the Composition of Foetal Tissue Grafts in a Rodent Model of Parkinson's Disease.Int J Mol Sci. 2022 Apr 22;23(9):4646. doi: 10.3390/ijms23094646. Int J Mol Sci. 2022. PMID: 35563037 Free PMC article.
-
Cell guidance ligands, receptors and complexes - orchestrating signalling in time and space.Curr Opin Struct Biol. 2020 Apr;61:79-85. doi: 10.1016/j.sbi.2019.11.007. Epub 2019 Dec 17. Curr Opin Struct Biol. 2020. PMID: 31862615 Free PMC article. Review.
-
Identification of Novel Genetic Loci Affecting Age at Onset of Parkinson's Disease: A Genome-wide Association Study.Mov Disord. 2025 Jan;40(1):77-86. doi: 10.1002/mds.30047. Epub 2024 Nov 6. Mov Disord. 2025. PMID: 39503264 Free PMC article.
-
Mechanistic advances in axon pathfinding.Curr Opin Cell Biol. 2020 Apr;63:11-19. doi: 10.1016/j.ceb.2019.12.003. Epub 2020 Jan 8. Curr Opin Cell Biol. 2020. PMID: 31927278 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous