

MiRNA-210 induces microglial activation and regulates microglia-mediated neuroinflammation in neonatal hypoxic-ischemic encephalopathy
- PMID: 31300734
- PMCID: PMC7608107
- DOI: 10.1038/s41423-019-0257-6
MiRNA-210 induces microglial activation and regulates microglia-mediated neuroinflammation in neonatal hypoxic-ischemic encephalopathy
Abstract
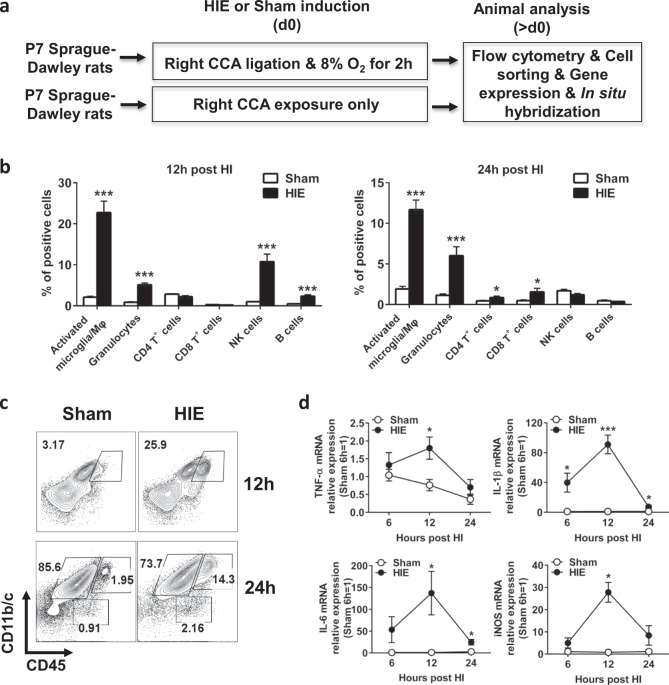
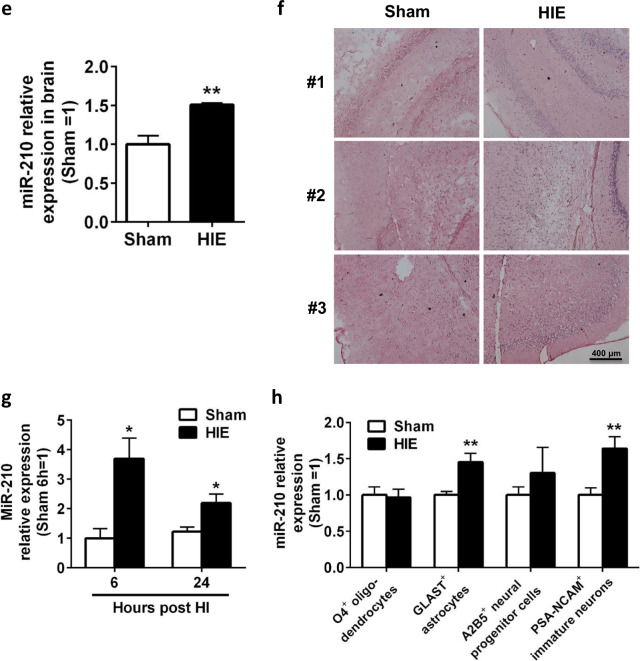
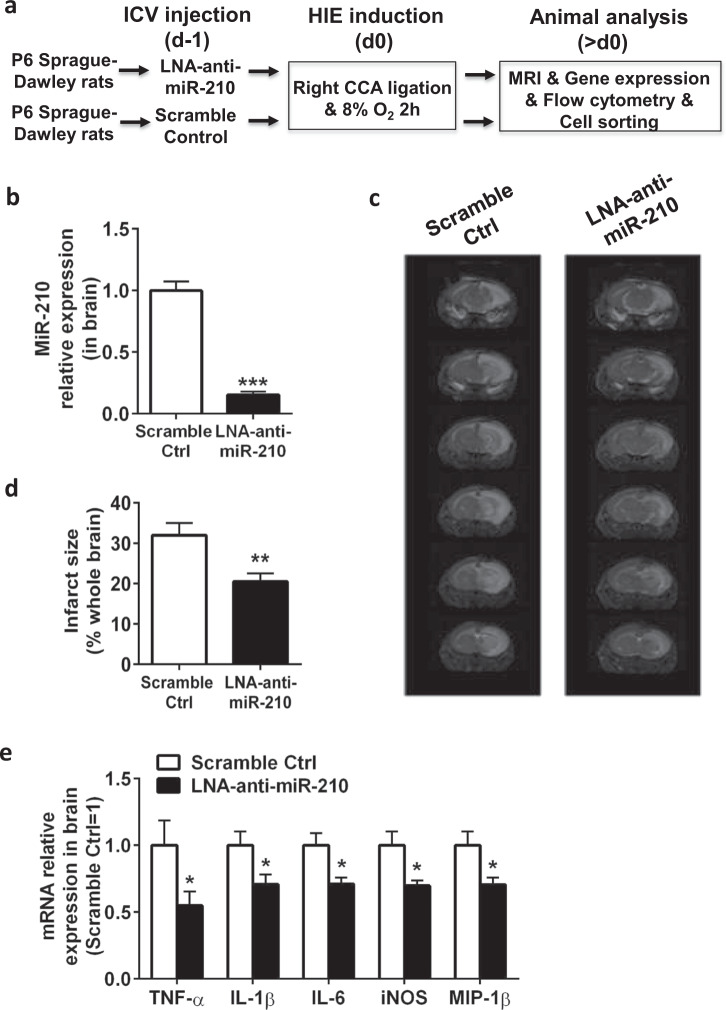
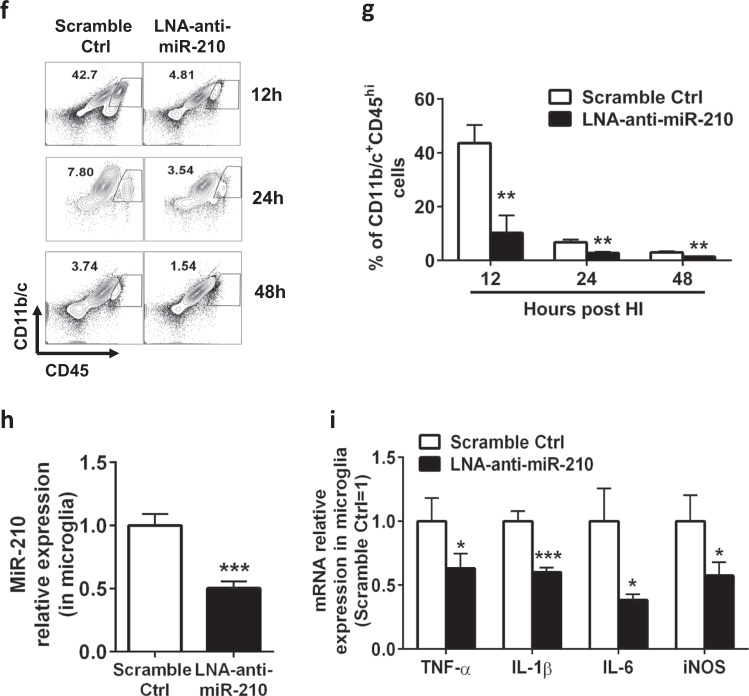
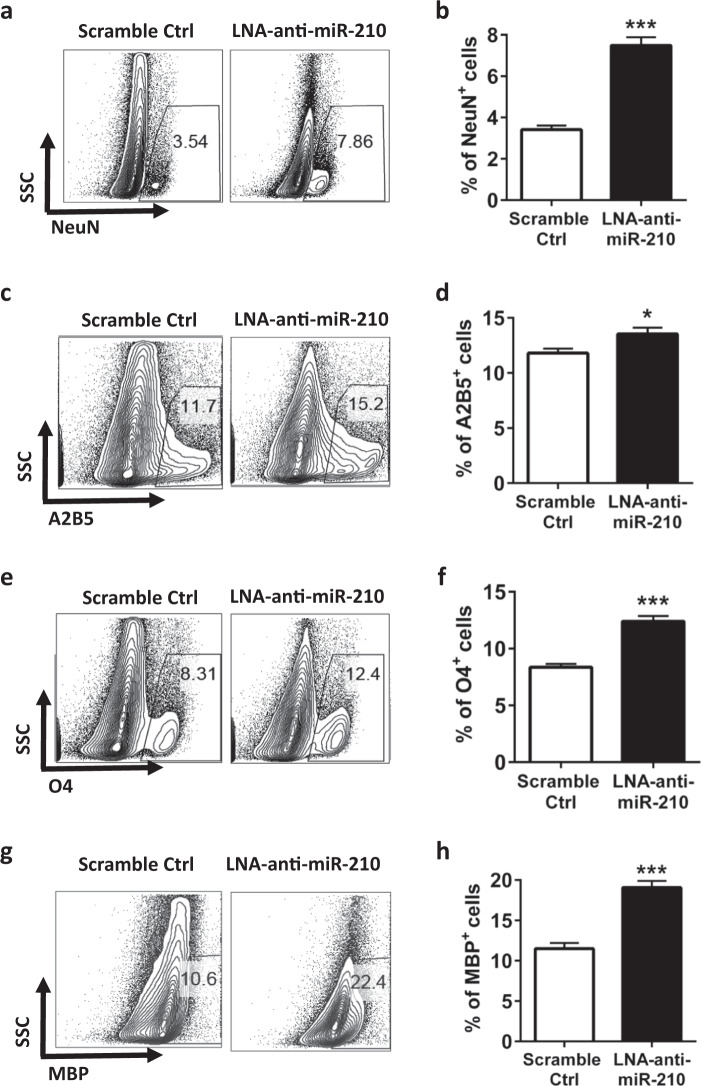
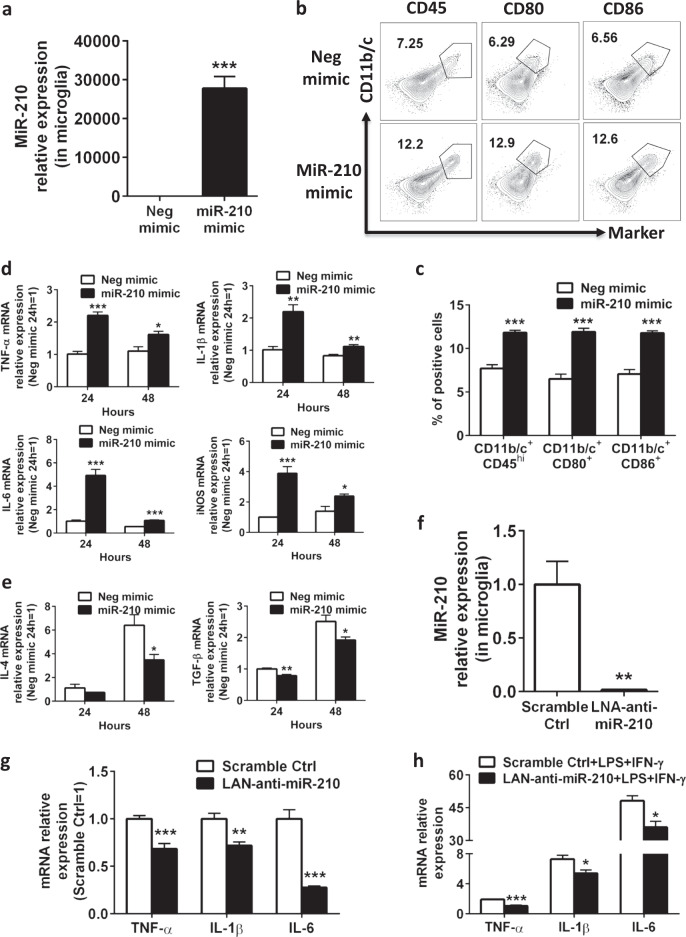
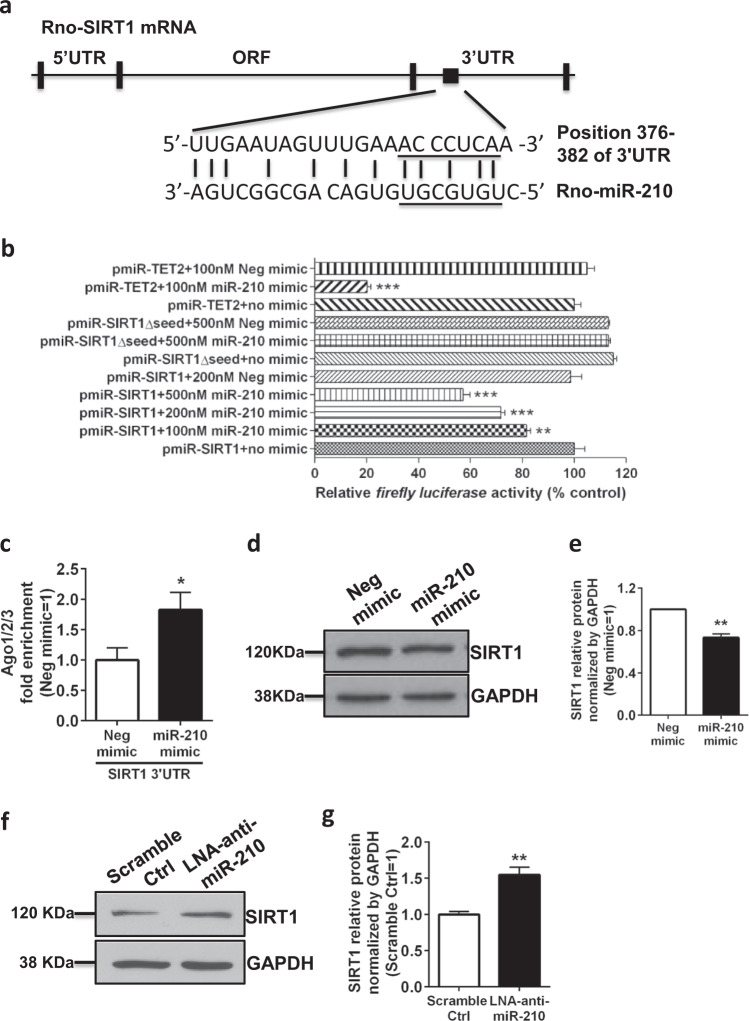
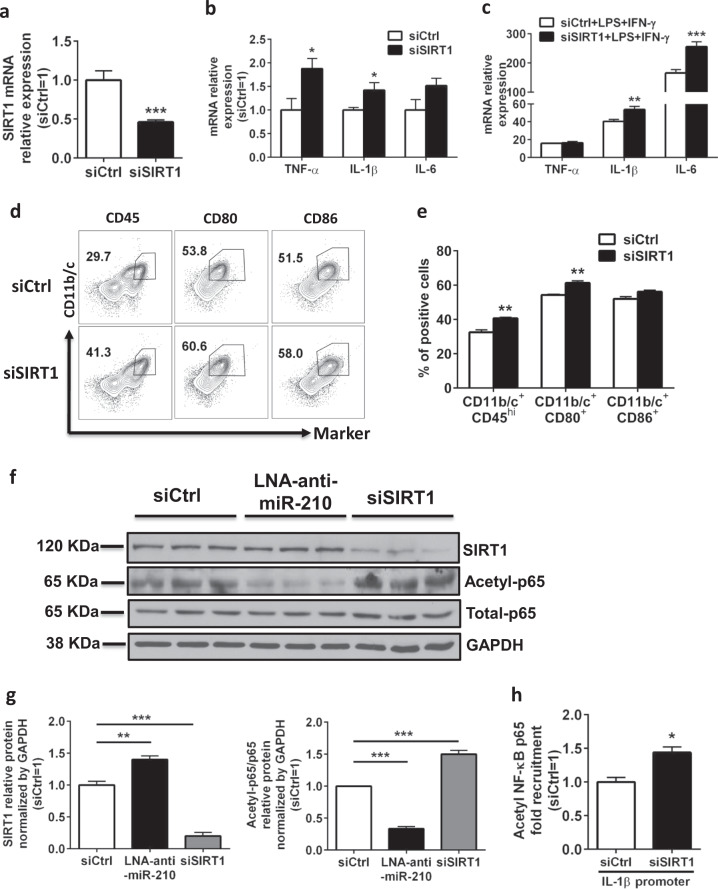
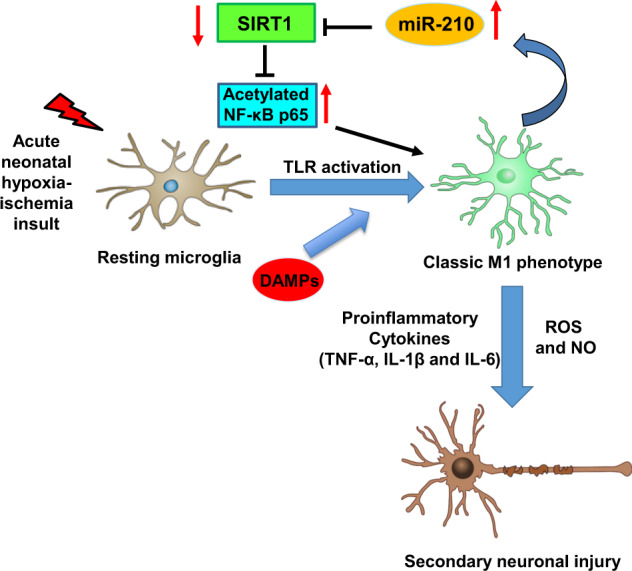
Neuroinflammation is a major contributor to secondary neuronal injury that accounts for a significant proportion of final brain cell loss in neonatal hypoxic-ischemic encephalopathy (HIE). However, the immunological mechanisms that underlie HIE remain unclear. MicroRNA-210 (miR-210) is the master "hypoxamir" and plays a key role in hypoxic-ischemic tissue damage. Herein, we report in an animal model of neonatal rats that HIE significantly upregulated miR-210 expression in microglia in the neonatal brain and strongly induced activated microglia. Intracerebroventricular administration of miR-210 antagomir effectively suppressed microglia-mediated neuroinflammation and significantly reduced brain injury caused by HIE. We demonstrated that miR-210 induced microglial M1 activation partly by targeting SIRT1, thereby reducing the deacetylation of the NF-κB subunit p65 and increasing NF-κB signaling activity. Thus, our study identified miR-210 as a novel regulator of microglial activation in neonatal HIE, highlighting a potential therapeutic target in the treatment of infants with hypoxic-ischemic brain injury.
Keywords: SIRT1; microRNA-210; microglial activation; neonatal hypoxic-ischemic encephalopathy; neuroinflammation.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Deng W. Neurobiology of injury to the developing brain. Nat. Rev. Neurol. 2010;6:328–336. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
