

Identification of endogenous small peptides involved in rice immunity through transcriptomics- and proteomics-based screening
- PMID: 31301098
- PMCID: PMC6953209
- DOI: 10.1111/pbi.13208
Identification of endogenous small peptides involved in rice immunity through transcriptomics- and proteomics-based screening
Abstract
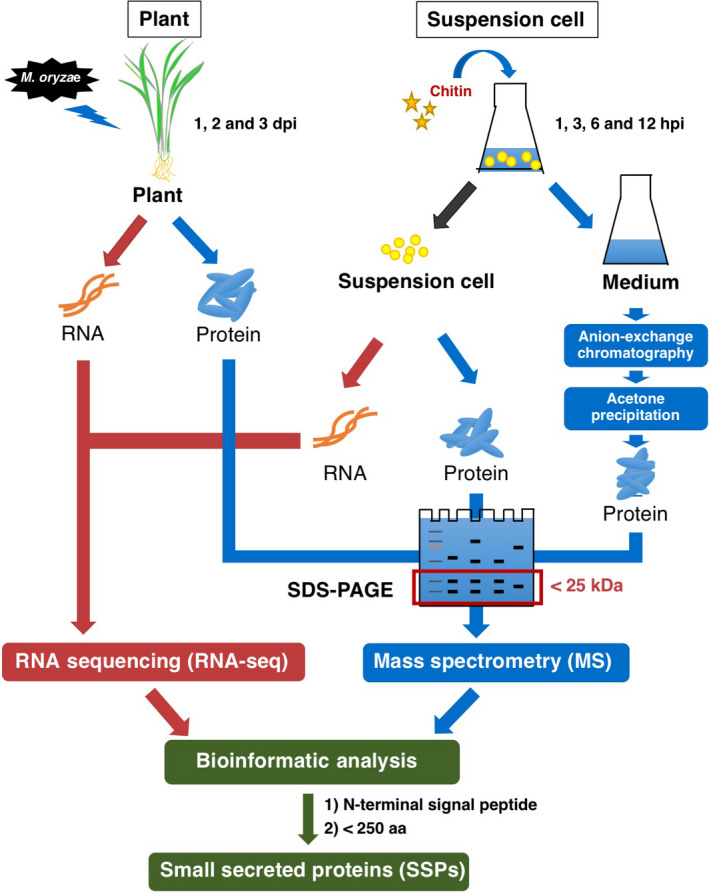
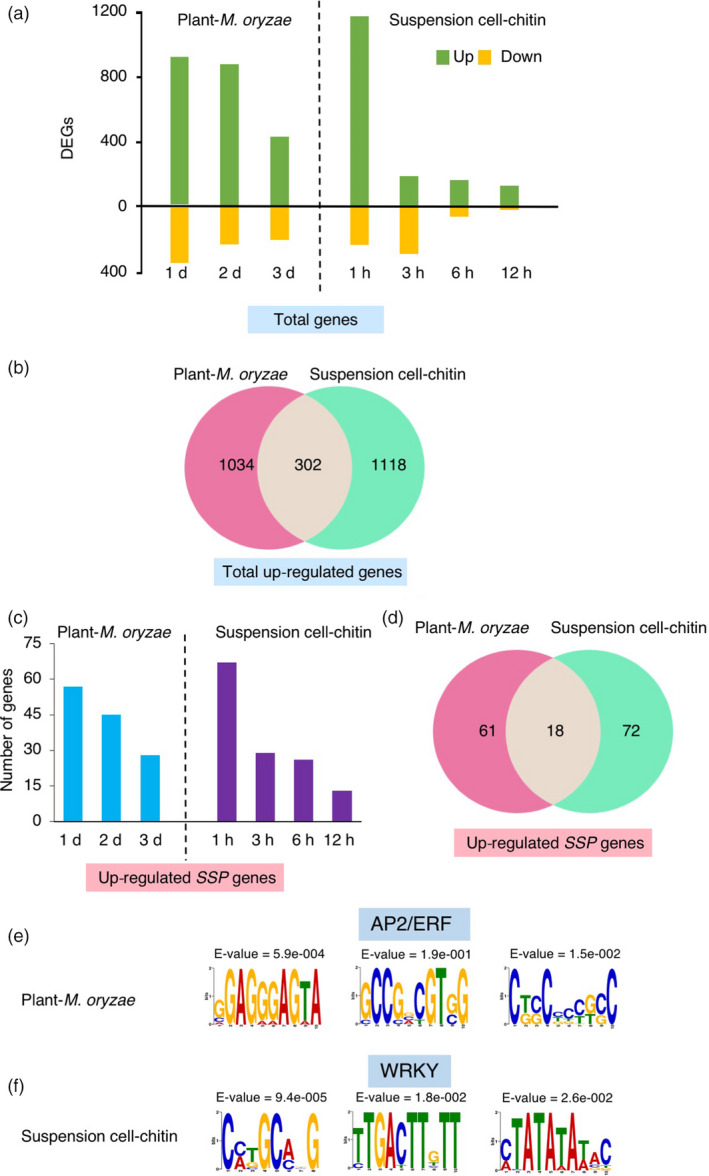
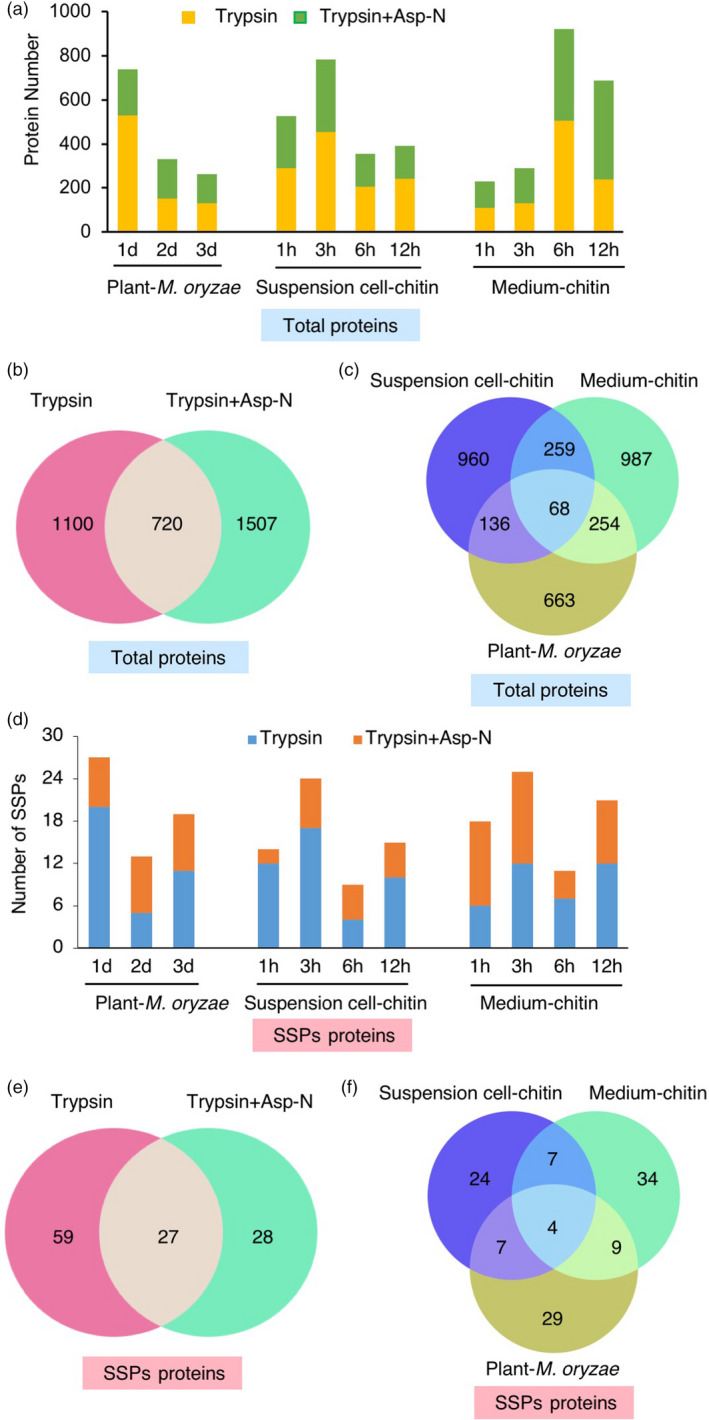
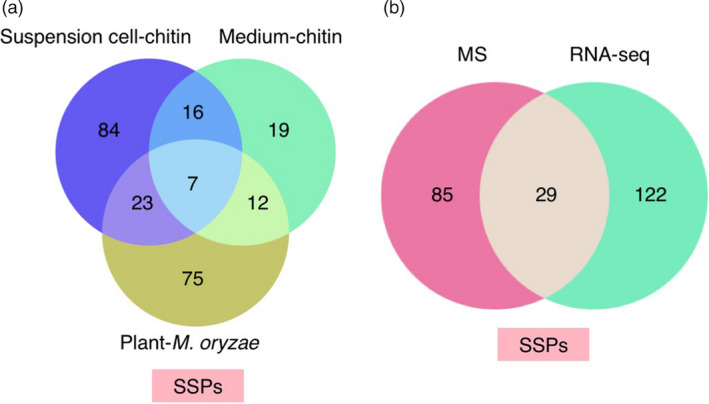
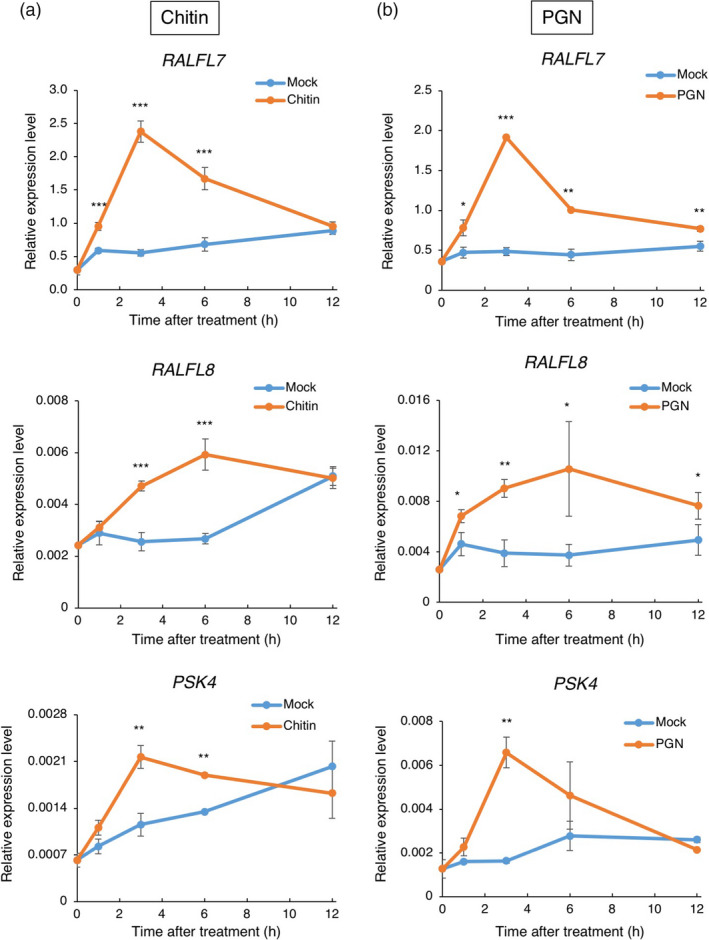
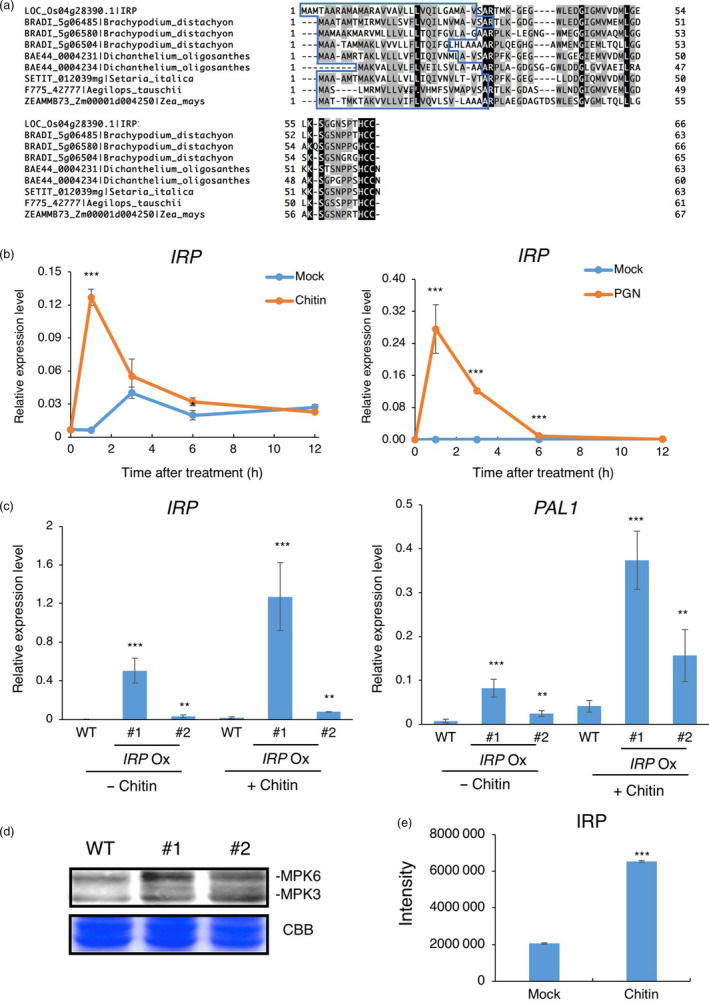
Small signalling peptides, generated from larger protein precursors, are important components to orchestrate various plant processes such as development and immune responses. However, small signalling peptides involved in plant immunity remain largely unknown. Here, we developed a pipeline using transcriptomics- and proteomics-based screening to identify putative precursors of small signalling peptides: small secreted proteins (SSPs) in rice, induced by rice blast fungus Magnaporthe oryzae and its elicitor, chitin. We identified 236 SSPs including members of two known small signalling peptide families, namely rapid alkalinization factors and phytosulfokines, as well as many other protein families that are known to be involved in immunity, such as proteinase inhibitors and pathogenesis-related protein families. We also isolated 52 unannotated SSPs and among them, we found one gene which we named immune response peptide (IRP) that appeared to encode the precursor of a small signalling peptide regulating rice immunity. In rice suspension cells, the expression of IRP was induced by bacterial peptidoglycan and fungal chitin. Overexpression of IRP enhanced the expression of a defence gene, PAL1 and induced the activation of the MAPKs in rice suspension cells. Moreover, the IRP protein level increased in suspension cell medium after chitin treatment. Collectively, we established a simple and efficient pipeline to discover SSP candidates that probably play important roles in rice immunity and identified 52 unannotated SSPs that may be useful for further elucidation of rice immunity. Our method can be applied to identify SSPs that are involved not only in immunity but also in other plant functions.
Keywords: Magnaporthe oryzae; immunity; proteomics; rice; signalling peptide; small secreted protein; transcriptomics.
© 2019 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Figures
References
-
- Akamatsu, A. , Wong, H.L. , Fujiwara, M. , Okuda, J. , Nishide, K. , Uno, K. , Imai, K. et al. (2013) An OsCEBiP/OsCERK1‐OsRacGEF1‐OsRac1 module is an essential early component of chitin‐induced rice immunity. Cell Host Microbe, 13, 465–476. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
