

N-Glycosylation influences human corticosteroid-binding globulin measurements
- PMID: 31307013
- PMCID: PMC6686952
- DOI: 10.1530/EC-19-0242
N-Glycosylation influences human corticosteroid-binding globulin measurements
Abstract
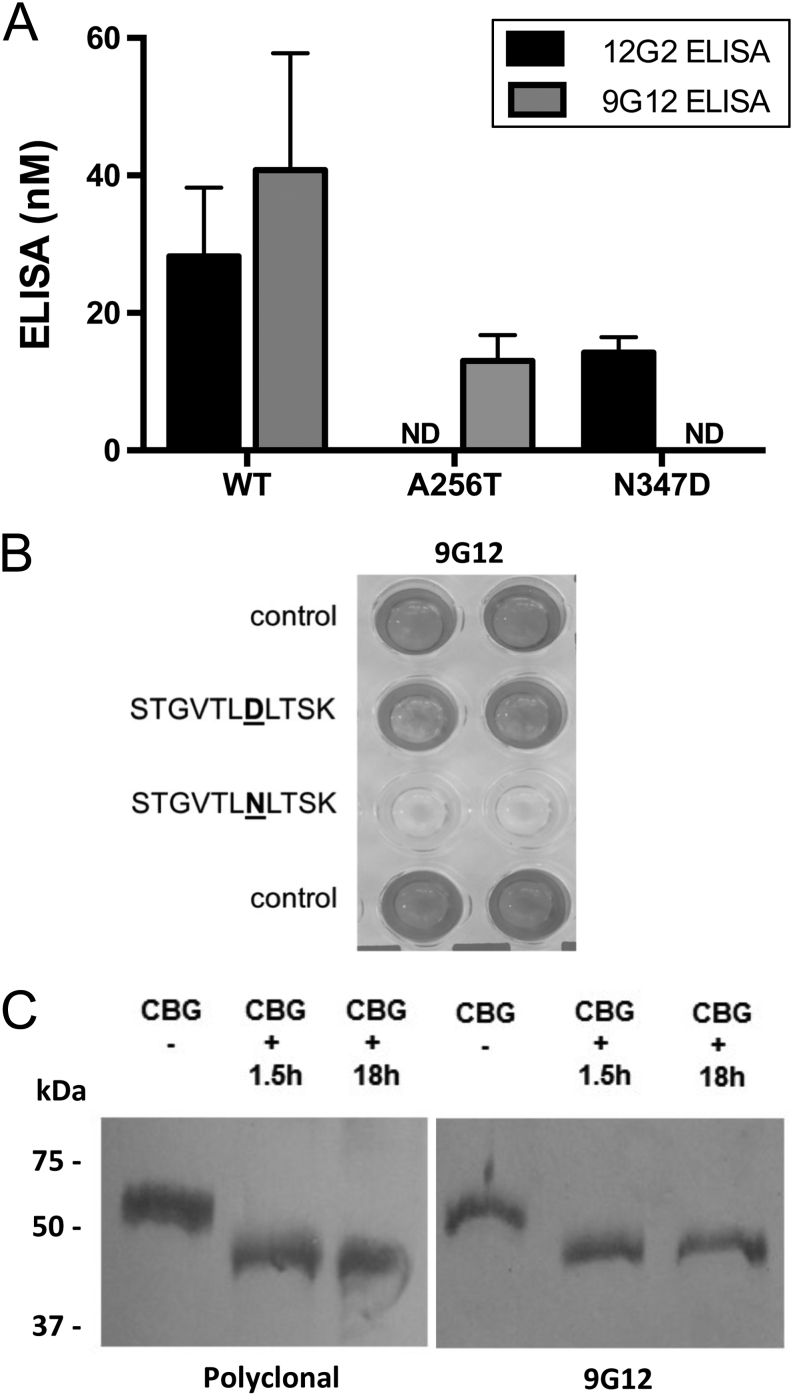
Objective: Discrepancies in ELISA measurements of human corticosteroid-binding globulin (CBG) using detection monoclonal antibodies that recognize an epitope (9G12) within its reactive center loop (RCL), versus an epitope (12G2) in a different location, have suggested that CBG with a proteolytically cleaved RCL exists in blood samples. We have previously been unable to verify this biochemically, and sought to determine if N-glycosylation differences account for discrepancies in ELISA measurements of CBG.
Methods and subjects: Molecular biological, biochemical and glycopeptide analyses were used to examine how N-glycosylation at specific sites, including at N347 within the RCL, affect CBG ELISA or steroid-binding capacity assay (BCA) measurements. Plasma from patients with congenital disorders of glycosylation (CDG) was also examined in these assays as examples of N-glycosylation defects.
Results: We demonstrate that an N-glycan at N347 within the CBG RCL limits the 9G12 antibody from recognizing its epitope, whereas the 12G2 antibody reactivity is unaffected, thereby contributing to discrepancies in ELISA measurements using these two antibodies. Qualitative differences in N-glycosylation at N238 also negatively affect the steroid-binding of CBG in the absence of an N-glycan at N347 caused by a T349A substitution. Desialylation increased both ELISA measurements relative to BCA values. Similarly, plasma CBG levels in both ELISAs were much higher than BCA values in several CDG patients.
Conclusions: Plasma CBG measurements are influenced by variations in N-glycosylation. This is important given the increasing number of CDG defects identified recently and because N-glycosylation abnormalities are common in patients with metabolic and liver diseases.
Keywords: congenital disorders of glycosylation; epitopes; glycosylation; monoclonal antibodies; protein structure; steroid binding.
Figures





References
-
- Hammond GL, Smith CL, Goping IS, Underhill DA, Harley MJ, Reventos J, Musto NA, Gunsalus GL, Bardin CW. Primary structure of human corticosteroid binding globulin, deduced from hepatic and pulmonary cDNAs, exhibits homology with serine protease inhibitors. PNAS 1987. 84 . ( 10.1073/pnas.84.15.5153) - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
